Действие радиации на иммунную систему. Влияние ионизирующего излучения на иммунную систему Иммунная реактивность и другие факторы
Читайте также
2.2 Влияние ионизирующей радиации на иммунитет
Малые дозы радиации, по-видимому, не оказывают заметного влияния на иммунитет. При облучении животных сублетальными и летальными дозами происходит резкое снижение резистентности организма к инфекции, что обусловлено рядом факторов, среди которых важнейшую роль играют: резкое повышение проницаемости биологических барьеров (кожи, дыхательных путей, желудочно-кишечного тракта и др.), угнетение бактерицидных свойств кожи, сыворотки крови и тканей, снижение концентрации лизоцима в слюне и крови, резкое уменьшение числа лейкоцитов в кровеносном русле, угнетение фагоцитарной системы, неблагоприятные изменения биологических свойств микробов, постоянно обитающих в организме, -- увеличение их биохимической активности, усиление патогенных свойств, повышение резистентности и др.
Облучение животных в сублетальных и летальных дозах приводит к тому, что из крупных микробных резервуаров (кишечник, дыхательные пути, кожа) в кровь и ткани поступает огромное количество бактерий.! При этом условно выделяют период стерильности (его продолжительность одни сутки), в течение которого микробов в тканях практически не обнаруживается; период обсемененности регионарных лимфатических узлов (обычно совпадает с латентным периодом); бактериемический период (длительность его 4--7 дней), который характеризуется появлением микробов в крови и тканях, и, наконец, период декомпенсации защитных механизмов, в течение которого отмечается резкое возрастание количества микробов в органах, тканях и крови (этот период наступает за несколько дней до гибели животных).
Под действием больших доз радиации, вызывающих частичную или полную гибель всех облученных животных, организм оказывается безоружным как к эндогенной (сапрофитной) микрофлоре, так и к экзогенным инфекциям. Считают, что в период разгара острой лучевой болезни и естественный, и искусственный иммунитет сильно ослаблен. Однако имеются данные, указывающие на более благоприятный исход течения острой лучевой болезни у животных, подвергшихся иммунизации до воздействия ионизирующего излучения. Вместе с тем экспериментально установлено, что вакцинация облученных животных отягощает течение острой лучевой болезни, и по этой причине она противопоказана до разрешения болезни. Напротив, через несколько недель после облучения в сублетальных дозах выработка антител постепенно восстанавливается, и поэтому уже через 1--2 мес после радиационного воздействия вакцинация вполне допустима.
Радиобиологи обладают весьма солидным багажом знаний о действии на биомакромолекулы, клетки, организмы высоких доз ионизирующего излучения, но не имеют ни достаточных данных...
Воздействие малых доз радиации
Огромное количество новых фактов, касающихся воздействия радиации, дали трагические последствия двух грандиозных радиационных катастроф: южно-уральской 1957 г. и чернобыльской 1986 г....
Воздействие малых доз радиации
Выдающийся шведский радиобиолог Р.М. Зиверт еще в 1950 г. пришел к заключению, что для действия радиации на живые организмы нет порогового уровня. Пороговый уровень - это такой...
Воздействие радиации на человека и окружающую среду
Считается, что радиация в любых дозах очень опасна. Ее влияние на живой организм может носить, как и позитивный характер: использование в медицине, так и негативный: лучевая болезнь. Любопытные результаты получили ученые...
Действие ионизирующего излучения на животных
В принципе все сельскохозяйственные животные, подвергшиеся действию ионизирующих излучений, могут быть разделены на две категории. К первой категории относятся животные, получившие летальные дозы радиации...
Естественный радиационный фон
Особенности воздействия радиации на живое вещество
Основную часть облучения население земного шара получает от естественных источников радиации. Большинство из них таковы, что избежать облучения от них совершенно невозможно...
Особенности воздействия радиации на живое вещество
В среднем примерно 2/3 эффективной эквивалентной дозы облучения, которую человек получает от естественных источников радиации, поступает от радиоактивных веществ, попавших в организм с пищей, водой и воздухом...
Особенности воздействия радиации на живое вещество
В своем последнем докладе НКДАР ООН впервые за 20 лет опубликовал подробный обзор сведений, относящихся к острому поражению организма человека, которое происходит при больших дозах облучения. Вообще говоря, радиация оказывает подобное действие...
Оценка экологической опасности осколков деления
Воздействие радиации на организм человека называют облучением. Во время этого процесса энергия радиации передается клеткам, тем самым разрушая их. Облучение может вызвать всевозможные заболевания: инфекционные осложнения...
Предельно допустимая концентрация вредных веществ
Предельно допустимый уровень (ПДУ) - это максимальный уровень воздействия радиации, шума, вибрации, магнитных полей и иных вредных физических воздействий, который не представляет опасности для здоровья человека, состояния животных, растений...
Солнечная радиация и ее влияние на природные и хозяйственные процессы
Цитология и охрана окружающей среды
Воздействие радиации на организм может быть различным, но почти всегда оно негативно. В малых дозах радиационное излучение может стать катализатором процессов, приводящих к раку или генетическим нарушениям...
лиферагивной активности (скорости деления) этих клеток, темпов деления и созревания в отделах морфологически распознаваемых предшественников. Показано, что после облучения темп пролиферации стволовых клеток усиливается, вследствие чего время удвоения их числа сокращается с 5-6 сут. до 1 суток.
Глубина и продолжительность аплазии кроветворной ткани оказывается тем больше, чем выше доза облучения. Однако из этого правила имеются исключения. После облучения в высоких дозах (у человека – 5,5 Гр) темп восстановления миелоидного ростка кроветворения оказывается существенно выше, и количество клеток в костном мозге и крови восстанавливается до контрольного уровня раньше, чем после воздействия в дозах 2-3 Гр, несмотря на то что степень опустошения в первом случае была гораздо сильнее. Причина этого явления не вполне ясна.
В случае неравномерного облучения или полного экранирования отдельных участков тела поражение органов кроветворной системы модифицируется за счет миграции клеток из менее облученных отделов. Этот механизм обеспечивает более быструю репопуляцию облученных органов, чем при общем равномерном облучении.
Стромальные элементы костного мозга довольно радиорезистентны. В первые недели после облучения в дозах, при которых развитие патологического процесса связано главным образом с поражением кроветворной системы, покоящиеся стромальные фибробласты мало повреждаются. Сохраняется, а иногда и возрастает их функциональная активность как продуцентов простагландина Е и колониестимулирующих факторов (КСФ). Поэтому при общем облучении в дозах, при которых сохраняется совместимое с выживанием число СКК, повреждения стромальных элементов мало выражены, и значение их в развитии аплазии костного мозга невелико. При облучении в более высоких дозах (порядка 100 Гр), применяемых, в частности, при лучевой терапии опухолей, часть зрелых фибробластов погибает, в то же время секреция ими простагландина Е и КСФ повышается.
Что касается родоначальных для фибробластов клеток (КОЕ-Ф), то они у человека достаточно радиочувствительны: D0 = 1,3 Гр. Кроме того, в периоде, когда костномозговое кроветворение восстанавливается, КОЕ-Ф пролиферируют очень вяло. Эти особенности могут иметь значение для развития отдаленных изменений в системе кроветворения, таких как апластическая анемия и т.п.
При морфологическом исследовании после облучения в высоких дозах (порядка 10 Гр) уже через несколько часов в красном костном мозге
наблюдают повреждения ядер клеток: пикноз, кариорексис (апоптоз), кариолизис. Иногда разрушается и цитоплазма (цитолиз). Костномозговые синусы расширяются, эритроциты выходят в паренхиму костного мозга. Еще через несколько часов начинается процесс фагоцитоза поврежденных миелокариоцитов: фрагменты ядер, а иногда и цельные клетки, обнаруживают в костномозговых макрофагах. У крыс уже через 1-2 сутки после сверхлетального облучения костный мозг очищается от продуктов клеточного распада, остаются лишь стромальные элементы. Развитие некротических изменений в клетках костного мозга сопровождается сосудистыми реакциями, и гистологическая картина костного мозга может быть охарактеризована как воспаление с характерными для него фазами альтерации, экссудации, а в последующем – развития репарационных процессов. В опустошенной отечной строме костного мозга наблюдаются геморрагии. Костный мозг через 3 суток после облучения в сверхлетальной дозе выглядит как «озеро крови». Незадолго перед смертью в костном мозге обнаруживаются бактерии.
При более низких дозах облучения изменения менее выражены. В случаях, заканчивающихся выздоровлением, вначале появляются отдельные очаги кроветворения, со временем они сливаются. У человека через 2 суток после облучения в дозах 2-6 Гр гистологическое исследование срезов костного мозга позволяет обнаружить снижение количества клеток эритроидного и гранулоцитарного ростков в 1,5-2 раза по сравнению с нормой, участки с обнаженной стромой, отечность основного вещества, мелкие кровоизлияния.
Позднее в костном мозге наблюдается прогрессирующее нарастание содержания стромальных элементов: фибробластов, ретикулярных клеток, макрофагов. После облучения в дозах 2-4 Гр в начале второй недели в период, соответствующий описанному ранее абортивному подъему, отмечается повышение митотической активности клеток костного мозга, увеличивается количество недифференцированных клеток. К концу третьей недели обнаруживается увеличение числа клеток гранулоцитарного и эритроидного ростков. При более высоких дозах облучения проявления абортивного подъема выражены значительно слабее.
С конца третьей недели после облучения в дозах более 2-3 Гр в костном мозге прогрессирует аплазия, проявляющаяся жировой атрофией. Площадь жировой ткани на срезах костного мозга может достигать 80%. Одновременно обнаруживаются и признаки регенерации: микроочаги недифференцированных клеток и клеток ранних генераций эритроидного и миелоидного ростков, а также усиление пролиферации стромальных клеток. В последующем очаги кроветворной ткани разрастаются
3.5. Действие ионизирующих излучений на критические системы организма
достаточно быстро, обеспечивая восстановление функции кроветворения.
Даже спустя год после облучения в дозах 2-4 Гр структура костного мозга восстанавливается не полностью. Сохраняются крупные очаги жировой дистрофии, увеличено по сравнению с нормой количество стромальных клеток, гемопоэтическая ткань в паренхиме костного мозга представлена не диффузно, а отдельными крупными очагами. После радиационного воздействия в дозах 5 Гр и выше в эти сроки в костном мозге наблюдаются распространенные очаги гипоплазии.
Морфологические изменения костномозговых клеток, связанные с их непосредственным повреждением (пикноз ядра, кариорексис, разрушение или отечность ядра), обычно удается зарегистрировать в первые часы после облучения. Разрушенные клетки довольно быстро удаляются. Позже начинают обнаруживаться клетки с аномалиями, связанными с нарушениями митозов: гигантские клетки, клетки с гиперсегментированным ядром, с аномальными митотическими фигурами (мосты в анаили телофазе), хромосомные фрагменты в интерфазных клетках. Максимум выхода этих клеток приходится на 12-24 ч после облучения. К третьему дню большинство таких клеток также исчезает.
3.5.3. Постлучевые изменения морфологического состава периферической крови
Убыль числа функциональных клеток начинается, когда последние клетки, поступившие к моменту облучения в созревающий отдел, выйдут в периферическую кровь, то есть через срок, соответствующий времени прохождения клеток через этот отдел в норме. Содержание в крови нейтрофильных гранулоцитов, прохождение которых через отдел созревания составляет 5-6 суток, начинает снижаться именно с этого времени.
Скорость уменьшения числа клеток в крови тем выше, чем короче продолжительность их циркуляции после выброса из костного мозга. Довольно большой срок между облучением и развитием максимальной нейтропении (это происходит к концу третьей недели) объясняется тем, что с середины второй недели после облучения в периферической крови развивается абортивный подъем содержания гранулоцитов.
Причины выброса в кровь в это время значительного числа зрелых нейтрофильных клеток связывают с влиянием на костный мозг экстрамедуллярных факторов, в частности, с возрастанием в крови на ран-

них сроках после облучения содержания катехоламинов и других биологически активных веществ.
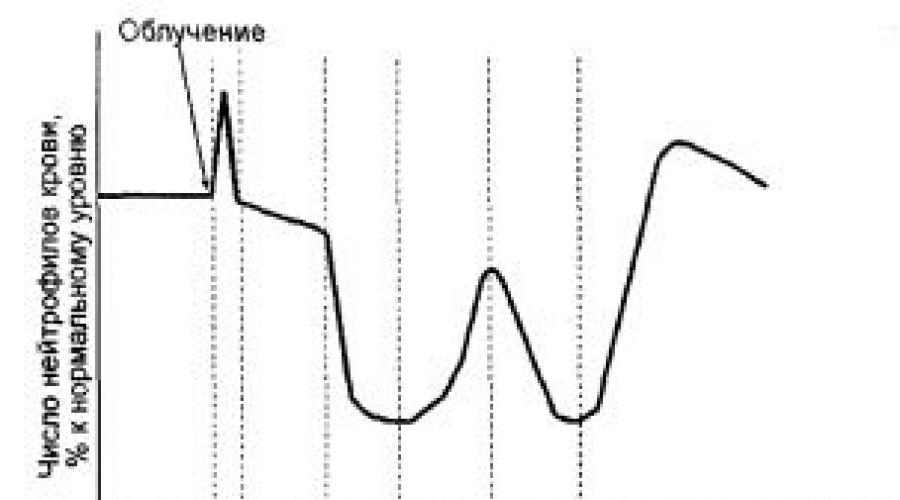
В динамике изменения числа нейтрофилов крови после облучения можно выделить несколько фаз:
− начальный или первичный нейтрофилез;
− лаг-фаза (в это время естественный выход в ткани нейтрофилов крови компенсирован поступлением этих клеток из пула созревания);
− первичное опустошение;
− абортивный подъем;
− вторичное опустошение;
− восстановление.
Лимфоциты погибают уже в ранние сроки после облучения; соответственно их уровень в периферической крови быстро снижается.
Рис. 11. Динамика изменений числа гранулоцитов в крови после общего облучения (по С. Киллмэну, 1974):
1 – первичный нейтрофилез; 2 – лаг-фаза; 3 – первичное опустошение; 4 – абортивный подъем; 5 – вторичное опустошение; 6 – восстановление
При резком снижении после облучения общего числа лейкоцитов в крови можно говорить о не менее глубоком уменьшении абсолютного количества моноцитов.
Содержание тканевых макрофагов в течение первой недели после облучения существенно не меняется. Функциональная активность этих клеток также не изменяется или даже оказывается повышенной. В то же время загруженность этих клеток продуктами тканевого распада снижает эффективность их участия в системе противомикробной защиты. В периоде выраженных клинических проявлений лучевого поражения число клеток системы мононуклеарных фагоцитов снижается.
3.5. Действие ионизирующих излучений на критические системы организма
Динамика содержания тромбоцитов после облучения имеет сходство с изменениями количества нейтрофилов. Абортивный подъем бывает выражен только после облучения в сравнительно невысоких (примерно до 3,5 Гр) дозах. В фазе вторичного опустошения наблюдается глубокая тромбоцитопения: к концу 3-4-й недели после облучения в среднелетальных дозах количество тромбоцитов достигает 5-8% от нормального уровня.
Содержание эритроцитов после облучения уменьшается в медленном темпе и умеренно, поскольку зрелые эритроциты достаточно радиорезистентны, а продолжительность жизни этих клеток составляет около 100 дней. Даже при полном прекращении их образования число эритроцитов в крови вследствие естественного отмирания снижается примерно на 1% за день (если нет осложняющих ситуацию кровотечений).
Глубина цитопении прямо зависит от дозы облучения. При дозах порядка 5-6 Гр нейтрофилы и тромбоциты могут вовсе не обнаруживаться в мазках периферической крови. Промежуток времени, необходимый для восстановления числа этих клеток до исходного уровня после облучения в высоких дозах, может оказаться, как об этом уже говорилось при характеристике изменений в кроветворных органах, короче, чем при воздействии меньших доз.
Кроме количественных сдвигов в клетках крови после облучения обнаруживаются и морфологические изменения: гомогенизация ядер лимфоцитов, появление лимфоцитов с микроядрами, гигантских гиперсегментированных нейтрофилов, гигантских тромбоцитов.
3.5.4. Влияние облучения на иммунную систему
Основной функцией иммунной системы является защита организма от воздействия чужеродных антигенов и контроль за поддержанием генетического постоянства внутренней среды организма. Эту функцию иммунная система выполняет с помощью естественных и адаптивных (приобретенных) механизмов. В основе естественного иммунитета лежит действие неспецифических механизмов, связанных с функционированием клеточных (нейтрофилы, макрофаги, ЕК-клетки (естественные киллеры) и др.) и гуморальных (комплемент, лизоцим, интерфероны и др.) факторов. Факторы естественного иммунитета относительно радиоустойчивы и поражаются лишь при очень высоких дозах облучения. Специфический иммунитет, основанный на свойствах
Т- и В-лимфоцитов избирательно отвечать на чужеродные вещества, напротив, высоко чувствителен к действию радиации.
Лимфоциты относятся к числу наиболее радиочувствительных клеток в организме, и гибель их отмечается уже после радиационного воздействия в десятых долях грея. При этом погибают не только молодые делящиеся клетки, но и (исключение из правила Бергонье и Трибондо) зрелые лимфоциты, которые в обычных условиях (без антигенной стимуляции) не делятся. Среди радиочувствительных клеток, подверженных интерфазной гибели при дозах, близких или даже более низких, чем вызывающие репродуктивную гибель стволовых кроветворных клеток, выделяются Т-лимфоциты (Т-хелперы и Т-супрессоры), В-лимфоциты и тимоциты вилочковой железы. Радиочувствительность В-лимфоцитов выше, чем Т-лимфоцитов, а радиационная устойчивость Т-супрессоров несколько больше, чем Т-хелперов. Тимоциты также различаются по своей радиочувствительности: максимальная радиопоражаемость отмечается у камбиальных клеток, а наибольшая радиорезистентность – у эпителиальных клеток. Кроме того, среди Т-лимфоцитов имеется сравнительно небольшая популяция радиоустойчивых клеток, которые сохраняют свою функциональную активность после облучения в весьма высоких дозах (6-10 Гр, а по некоторым данным – до 20 Гр). Эти клетки являются одновременно кортизонорезистентными. Их содержание составляет около 3-8% от всех Т-лимфоцитов, и, возможно, они представляют собой Т-клетки памяти.
С высокой радиочувствительностью зрелых популяций лимфоцитов крови и их интерфазной гибелью в первый день после воздействия ионизирующего излучения связано и быстрое развитие лимфопении после облучения. Интерфазная гибель лимфоцитов не связана с моментом клеточного митоза, она начинается через 6 часов и завершается через 3 суток после радиационного воздействия. Разрушение лимфоцитов после облучения происходит как в лимфоидных органах (тимус, лимфатические узлы, селезенка, лимфоидные образования в кишечнике), так и в периферической крови и лимфе. В результате можно обнаружить снижение числа лимфоцитов в крови уже через десятки минут после облучения, а к 3-м суткам число лимфоцитов снижается до минимальных величин. Глубина снижения уровня лимфоцитов, так же как и других клеток периферической крови, прямо зависит от дозы облучения. Следует подчеркнуть, что послерадиационное падение числа лимфоцитов, наряду с гранулоцитопенией, является основной причиной развития синдрома инфекционных осложнений в периоде разгара острой лучевой болезни.
3.5. Действие ионизирующих излучений на критические системы организма
Таблица 37 – Радиочувствительность некоторых клеток иммунной системы человека
(по данным А.А. Ярилина, 1989, 1997; Т. Szepesi, Т.М. Fliedner, 1989)
Вид клеток |
Д0 , Гр |
Полипотентные стволовые кроветворные клетки |
|
Клетки-предшественники гранулоцитов |
|
Нейтрофилы крови |
|
Клетки-предшественники моноцитов |
|
Моноциты крови |
|
ЕК-клетки (естественные киллеры) |
|
Клетки-предшественники лимфоцитов: ранние стадии |
|
поздние стадии |
|
Лимфоциты крови: Т-лимфоциты |
|
В-лимфоциты |
|
Клетки-предшественники тимоцитов – камбиальные клетки тимуса |
|
Эпителиальные клетки тимуса |
|
Дендритные клетки тимуса |
|
Воздействие радиации на лимфоидную ткань приводит не только к гибели лимфоцитов, но и вызывает значительные изменения их функциональной активности. Это, в свою очередь, может приводить к извращению иммунного ответа как в ближайшие сроки, так и (что особенно важно) в отдаленном периоде после лучевого воздействия.
Так, уже в ближайшие минуты-часы после облучения как для В-лим- фоцитов, так и особенно для Т-лимфоцитов, характерна утрата имеющихся на их поверхности клеточных рецепторов для различных антигенов, что связано с вмешательством ионизирующих излучений в процесс реаранжировки генов TCR (антигенраспознающего рецептора Т-лим- фоцитов). Пострадиационное изменение экспрессии молекул адгезии приводит к нарушению распределения лимфоцитов в крови и лимфоидных органах и фактически искажает пространственную организацию иммунной системы.
Уже в ранние сроки после облучения значительно снижается способность В-лимфоцитов к выработке специфических иммуноглобулинов в ответ на антигенную стимуляцию. Это угнетение прямо коррелирует с динамикой депопуляции лимфоидных органов, а наиболее выражено в случае введения антигена через 1-2 сут. после облучения. При введении антигена незадолго до облучения выработка антител может даже увеличиваться. В случае предварительной иммунизации перед воз-
действием радиации «вторичный ответ» на повторное введение антигена после облучения существенно не нарушается.
Еще одним ближайшим результатом радиационного воздействия является снижение пролиферативной активности Т-лимфоцитов, их миграционных свойств, способности инактивировать несингенные КОЕ. Гибель в результате облучения Т-лимфоцитов сопровождается снижением в организме их цитотоксических функций, что проявляется подавлением некоторых реакций гиперчувствительности замедленного типа, реакций типа «трансплантат против хозяина» и др. Выраженность угнетения этих реакций зависит в значительной мере и от уровня функциональной активности Т-лимфоцитов, которая оказывается подавленной уже после облучения в дозах 0,15-0,20 Гр.
Воздействие ионизирующих излучений приводит к нарушению баланса Т-хелперов классов Th1 и Th2, которые определяют соотношение клеточной опосредованной и гуморальной составляющей иммунного ответа, а также ряда проявлений иммунопатологии. Напомним, что продукты Th2 – интерлейкин-2, γ-интерферон, β-туморнекротический фактор – обеспечивают развитие клеточного иммунитета, а продукты Th2 – интерлейкин-4, -5, -10 – служат медиаторами гуморального ответа. Эти клетки находятся в соотношениях антагонизма, реализуемого с участием γ-интерферона и интерлейкина-10.
Дифференцировка Thl поддерживается макрофагами через выработку интерлейкина-12, а развитие Th2 регулируется интерлейкином-4. В свою очередь продукт Thl γ-интерферон стимулирует активность макрофагов.
3.5. Действие ионизирующих излучений на критические системы организма
Установлено, что чем выше зависимость иммунного ответа от тимуса, тем сильнее проявляется эффект радиации. Действие ионизирующих излучений на тимусзависимое звено иммунной системы складывается из непосредственного влияния на Т-клетки и опосредованного действия через строму тимуса. Активность стромы тимуса в ранние сроки после облучения может повышаться, а в более поздние, как правило, подавляется, что сопровождается ускоренной передачей полномочий тимуса периферическому отделу иммунной системы и развитием проявлений иммунологического старения.
В раннем пострадиационном периоде повышается и вероятность развития аутоиммунных реакций, выраженность которых нарастает с увеличением дозы облучения. Однако аутоиммунные процессы проявляются и в поздние сроки после лучевого воздействия, а также при действии малых доз радиации. Ряд исследователей полагают, что для малых доз и интенсивностей ионизирующих излучений развитие аутоиммунных процессов даже более характерно, чем для последствий облучения в высоких дозах.
При действии высоких доз ионизирующих излучений, убивающих большую часть лимфоцитов, нарушается формирование антигенраспознающего механизма. Клетки, обусловливающие селекцию тимоцитов, сильно различаются по своей радиочувствительности: эпителиальные клетки устойчивы к действию радиации в дозах до 8-10 Гр, а дендритные клетки погибают уже при дозах 2-4 Гр. В связи с этим процесс положительной селекции является относительно радиоустойчивым, а облучение в малых дозах даже может повышать его эффективность. Напротив, процесс отрицательной селекции нарушается уже при действии относительно малых доз радиации, вследствие чего часть аутореактивных клонов может сохраниться, и в последующем стать источником аутоагрессии. В поздние сроки после облучения могут страдать не только дендритные, но и эпителиальные клетки тимуса. Это связано с гибелью их относительно радиочувствительных предшественников – делящихся камбиальных клеток (D0 для них составляет 2,5-3,7 Гр). В результате этого снижается число прошедших дифференцировку Т-лимфоцитов, уменьшается общая численность тимоцитов (подобный процесс наблюдается при старении) и, как следствие, возрастает вероятность развития аутоиммунных и опухолевых процессов.
Еще одним фактором, приводящим к прогрессированию аутоиммунных процессов в облученном организме, является ранняя пострадиационная гибель специальной популяции супрессорных клеток, сдерживающих образование В1-клетками естественных аутоантител к
эндогенным субстанциям. Устранение этих клеток облучением, а они погибают уже при дозах 4-6 Гр, приводит к усилению выработки естественных аутоантител и, как следствие, развитию органоспецифических аутоиммунных процессов.
Важным аспектом действия ионизирующих излучений на иммунитет является также их влияние на систему цитокинов – продуктов активированных клеток иммунной системы, играющих ключевую роль в регуляции гемопоэза и межклеточного взаимодействия в ходе развития воспаления и иммунного ответа. Действие радиации на эту систему сильно зависит от природы клеток-продуцентов цитокинов. Так, образование лимфокинов in vivo подавляется в связи с массовой гибелью вырабатывающих их лимфоцитов, хотя сам по себе процесс продукции цитокина может стимулироваться радиацией (как это имеет место в случае с интерлейкином-2). В то же время воздействие ионизирующих излучений приводит к увеличению выработки интерлейкина-1, -6 и фактора некроза опухолей, макрофагами, стромальными и эпителиальными клетками тимуса. Стимуляция выработки этих цитокинов облучением особенно интересна, потому что интерлейкин-1 и фактор некроза опухолей, сами по себе обладают радиопротекторным действием, реализуемым при участии интерлейкина-6, а в сочетании с интерлейкином-1 радиозащитная активность проявляется также у гранулоцитарного и гранулоцитарно-макрофагального факторов. Эти факты, вероятно, свидетельствуют о том, что некоторые эффекты радиации направлены на ослабление или ликвидацию вызванных ею же последствий.
Таким образом, ионизирующие излучения существенным образом влияют на иммунную систему, вызывая широкий спектр ее реакций – от изменения регуляции иммунного ответа до гибели иммунокомпетентных клеток. Так, изменение экспрессии молекул адгезии, приводя к нарушениям распределения лимфоцитов, искажает пространственную организацию иммунной системы. Ее временная организация нарушается вследствие вмешательства радиации в процесс реаранжировки генов TCR, повреждения эпителия тимуса и связанного с этим «перевода иммунологических часов» в сторону старения.
3.5.5. Действие ионизирующей радиации на желудочно-кишечный тракт
В желудочно-кишечном тракте наиболее радиочувствительной является тонкая кишка, D0 для стволовых клеток эпителия тонкой кишки составляет около 1 Гр, тогда как в толстой кишке этот показатель до-
Источники ионизирующего излучения (радионуклиды) могут находиться вне организма и (или) внутри его. Если животные подвергаются воздействию излучения извне, то говорят о внешнем облучении, а воздействие ионизирующих излучений на органы и ткани от инкорпорированных радионуклидов называют внутренним облучением. В реальных условиях чаще всего возможны различные варианты и внешнего, и внутреннего облучения. Такие варианты воздействия называются сочетанными радиационными поражениями.
Доза внешнего облучения формируется главным образом за счет воздействия г-излучения; б- и в-излучения не вносят существенного вклада в общее внешнее облучение животных, так как они в основном поглощаются воздухом или эпидермисом кожи. Радиационное поражение кожных покровов в-частицами возможно в основном при содержании скота на открытой местности в момент выпадения радиоактивных продуктов ядерного взрыва или других радиоактивных осадков.
Характер внешнего облучения животных во времени может быть различным. Возможны разные варианты однократного облучения, когда животные подвергаются радиационному воздействию в течение короткого промежутка времени. В радиобиологии принято считать однократным облучением воздействие радиации на протяжении не более 4 сут. Во всех случаях, когда животные подвергаются внешнему облучению с перерывами (они могут быть различными по продолжительности), имеет место фракционированное (прерывистое) облучение. При непрерывном длительном воздействии ионизирующего излучения на организм животных говорят о пролонгированном облучении.
Выделяют общее (тотальное) облучение, при котором радиационному воздействию подвергается все тело. Этот вид облучения имеет место, например, при обитании животных на территории, загрязненной радиоактивными веществами. Кроме того, в условиях специальных радиобиологических исследований может осуществляться местное облучение, когда радиационному воздействию подвергается та или иная часть тела! При одной и той же дозе облучения наиболее тяжелые последствия наблюдаются при общем облучении. Например, при облучении всего тела животных в дозе 1500 Р отмечается практически 100%-ная их гибель, тогда как облучение ограниченного участка тела (головы, конечностей, щитовидной железы и т. д.) каких-либо серьезных последствий не вызывает. В дальнейшем рассматриваются последствия только общего внешнего облучения животных.
Влияние ионизирующей радиации на иммунитет
Малые дозы радиации, по-видимому, не оказывают заметного влияния на иммунитет. При облучении животных сублетальными и летальными дозами происходит резкое снижение резистентности организма к инфекции, что обусловлено рядом факторов, среди которых важнейшую роль играют: резкое повышение проницаемости биологических барьеров (кожи, дыхательных путей, желудочно-кишечного тракта и др.), угнетение бактерицидных свойств кожи, сыворотки крови и тканей, снижение концентрации лизоцима в слюне и крови, резкое уменьшение числа лейкоцитов в кровеносном русле, угнетение фагоцитарной системы, неблагоприятные изменения биологических свойств микробов, постоянно обитающих в организме, -- увеличение их биохимической активности, усиление патогенных свойств, повышение резистентности и др.
Облучение животных в сублетальных и летальных дозах приводит к тому, что из крупных микробных резервуаров (кишечник, дыхательные пути, кожа) в кровь и ткани поступает огромное количество бактерий.! При этом условно выделяют период стерильности (его продолжительность одни сутки), в течение которого микробов в тканях практически не обнаруживается; период обсемененности регионарных лимфатических узлов (обычно совпадает с латентным периодом); бактериемический период (длительность его 4--7 дней), который характеризуется появлением микробов в крови и тканях, и, наконец, период декомпенсации защитных механизмов, в течение которого отмечается резкое возрастание количества микробов в органах, тканях и крови (этот период наступает за несколько дней до гибели животных).
Под действием больших доз радиации, вызывающих частичную или полную гибель всех облученных животных, организм оказывается безоружным как к эндогенной (сапрофитной) микрофлоре, так и к экзогенным инфекциям. Считают, что в период разгара острой лучевой болезни и естественный, и искусственный иммунитет сильно ослаблен. Однако имеются данные, указывающие на более благоприятный исход течения острой лучевой болезни у животных, подвергшихся иммунизации до воздействия ионизирующего излучения. Вместе с тем экспериментально установлено, что вакцинация облученных животных отягощает течение острой лучевой болезни, и по этой причине она противопоказана до разрешения болезни. Напротив, через несколько недель после облучения в сублетальных дозах выработка антител постепенно восстанавливается, и поэтому уже через 1--2 мес после радиационного воздействия вакцинация вполне допустима.
Введение Двадцатому веку последовательно приписывали три наименования - атомный, космический и век биологии. Можно смело утверждать, что первое из них пока наиболее емко, ибо есть все основания полагать, что успехи в познании тайн атомного ядра и управление его энергией окажут решающее влияние на все проблемы жизни на нашей планете и за ее пределами. Явление радиоактивности было открыто около ста лет назад Пьером Кюри и Марией Склодовской-Кюри. Именно это открытие положило начало бурному развитию новых направле
ний в химии и физике, которые, в свою очередь, стали фундаментом для создания атомно-промышленного комплекса. Первые предприятия атомной промышленности были направлены на создание атомной бомбы, что и было впервые сделано в США. В боевых целях ядерное оружие было применено 6 и 9 августа 1945 года, когда американцами были взорваны две атомные бомбы над японскими городами Хиросима и Нагасаки. Первым предприятием атомной промышленности, созданным в СССР, стало производственное объед
инение «Маяк», предназначенное для получения делящихся ядерных материалов. Первые предприятия ядерного комплекса формировались в условиях «гонки вооружения», к тому же эффекты воздействия радиации на организм человека и окружающую среду были мало изучены, что и привело к необдуманному сбросу отходов, крупномасштабному загрязнению окружающей среды и росту числа заболеваний у работников атомной промышленности и населения, проживающего в зоне радиоактивного загрязнения, вследствие неверного нормирования доз облучени
я. В настоящее время атомно-промышленный комплекс представляет собой разветвленную сеть предприятий с различными целями и задачами. В него входят предприятия военно-промышленного комплекса, АЭС, научно-исследовательские центры и институты. За последние десятилетия произошла переоценка эффектов влияния атомной радиации на человека и окружающую среду. Был введен запрет на испытания и распространение ядерного оружия, а также подписано несколько договоров о сокращении ядерного вооружения.
29 июля 1957 года была учреждена МАГАТЭ – автономная межправительственная организация по вопросам мирного использования ядерной энергии. Целью ее создания стал контроль за деятельностью стран с развитой атомной промышленностью в соответствии с целями и принципами ООН, направленными на укрепление мира и поощрение международного сотрудничества. Международные организации, работающие в сфере изучения влияния радиации на человека и ОС, периодически пересматривали степень ее опасности в сторону повышения. С 30-ых годов этот у
ровень возрос в тысячу раз. Международная комиссия радиационной защиты официально признала концепцию беспорогового действия радиации на здоровье человека. Однако и в настоящее время научные дискуссии о механизмах воздействия ионизирующего излучения и его отдаленных последствий на живой организм не завершены, и многие вопросы требуют дальнейшего изучения. Исследования в области данной проблемы по-прежнему остаются актуальными как из-за постоянного имеющегося риска радиоактивного заражения окружающей среды, так и из
За риска потерь здоровья лиц, уже подвергшихся радиационному воздействию. Виды ионизирующих излучений На состояние окружающей среды и на живые организмы оказывают сильное влияние различные экологические факторы. Экологический фактор - любое условие среды, способное оказывать прямое или косвенное воздействие на живые организмы. Экологические факторы делятся на три категории: 1. абиотические - факторы неживой природы, 2. биотические - факторы живой природы и 3. антропогенные - факторы человеческой деятельност
и. Важным абиотическим фактором наземной среды является ионизирующее излучение - это излучение с очень высокой энергией, способное выбивать электроны из атомов и присоединять их к другим атомам с образованием пар положительных и отрицательных ионов. Существует два типа ионизирующих излучений: корпускулярное, состоящее из частиц с массой покоя, отличной от нуля (альфа, бета и нейтронное излучение), и электромагнитное (гамма- и рентгеновское излучение) с очень малой длиной волны. Альфа-излучение представляет собой поток ядер
гелия, обладающих большой скоростью. Эти ядра имеют массу 4 и заряд +2. Они образуются при радиоактивном распаде ядер и при ядерных реакциях. Энергия альфа-частиц не превышает нескольких МэВ (1 эВ=1,60206*10-19 Дж). Длина пробега альфа-частиц в воздухе обычно менее 10 см (под длиной пробега частицы понимается наибольшее расстояние от источника излучения, при котором еще можно обнаружить частицу, до ее поглощения веществом). В воде или в мягких тканях человеческого те
ла, плотность которых более чем в 700 раз превышает плотность воздуха, длина пробега альфа-частиц составляет несколько десятков микрометров. За счет своей большой массы при взаимодействии с веществом альфа-частицы быстро теряют свою энергию. Это объясняет их низкую пронизывающую способность и высокую удельную ионизацию: при движении в воздушной среде альфа-частица на 1 см своего пути образует несколько десятков тысяч пар заряженных частиц - ионов. Бета-излучение представляет собой поток электронов (β-
излучение) или позитронов (+-излучение), возникающих при радиоактивном распаде. Масса бета-частиц в несколько десятков тысяч раз меньше массы альфа-частиц. В зависимости от природы источника бета-излучений скорость этих частиц может лежать в пределах 0,3-0,99 скорости света. Энергия бета-частиц не превышает нескольких МэВ, длина пробега в воздухе составляет приблизительно 1800 см, а в мягких тканях человеческого тела -
2,5 см. Проникающая способность бета-частиц выше, чем альфа-частиц (из-за меньших массы и заряда). Нейтронное излучение представляет собой поток ядерных частиц, не имеющих электрического заряда. Масса нейтрона приблизительно в 4 раза меньше массы альфа-частиц. В зависимости от энергии различают медленные нейтроны (с энергией 1 КэВ), нейтроны промежуточных энергий (от 1 до 500 КэВ) и быстрые нейтроны (от 5
00 КэВ до 20 МэВ). При неупругом взаимодействии нейтронов с ядрами атомов среды возникает вторичное излучение, состоящее из заряженных частиц и гамма-квантов (гамма-излучение). При упругих взаимодействиях нейтронов с ядрами может наблюдаться обычная ионизация вещества. Проникающая способность нейтронов зависит от их энергии, но она существенно выше, чем у альфа- или бета-частиц. Так, длина пробега нейтронов промежуточных энергий составляет около 15 м в воздушной среде и 3 см
в биологической ткани, аналогичные показатели для быстрых нейтронов - соответственно 120 м и 10 см. Таким образом, нейтронное излучение обладает высокой проникающей способностью и представляет для человека наибольшую опасность из всех видов корпускулярного излучения. Мощность нейтронного потока измеряется плотностью потока нейтронов (нейтрон.см2*с). Гамма-излучение (γ-излучение) представляет собой электромагнитное излучение с высокой энергией и с малой длиной волны (порядка 3*
10-2 нм). Оно испускается при ядерных превращениях или взаимодействии частиц. Высокая длина (0,01-3 МэВ) и малая длина волны обуславливает большую проникающую способность гамма-излучения. Гамма-лучи не отклоняются в электрических и магнитных полях. Это излучение обладает меньшей ионизирующей способностью, чем альфа- и бета- излучения. Рентгеновское излучение может быть получено в специальных рентгеновских трубах, в ускорителях электронов, в среде, окружающей источник бета-излучения, и др. Рентгеновские лучи предста
вляют собой один из видов электромагнитного излучения. Энергия его обычно не превышает 1 МэВ. Рентгеновское излучение, как и гамма-излучение, обладает малой ионизирующей способностью и большой скоростью. При распаде ядер атомов его продукты вылетают с большой скоростью. Встречая на своем пути ту или иную преграду, они производят в ее веществе различные изменения. Воздействие излучения на вещество будет тем больше, чем больше распадов происходит в единицу времени. Д
ля характеристики числа распадов вводится понятие активности (А) радиоактивного вещества, под которым понимают число самопроизвольных ядерных превращений dN в этом веществе за малый промежуток времени dt, деленное на этот промежуток времени: A = dN / dt. Влияние ионизирующего излучения на вещество характеризуется поглощенной дозой – количеством энергии, переданным единице массы вещества. В системе СИ единицей поглощенной дозы служит грей (
Гр) – доза, при которой 1кг вещества передается энергия 1Дж. Иногда используют внесистемную единицу – рад: 1рад=100эрг/г=10-2Гр. Поглощенная доза ионизирующего излучения является основной физической величиной, определяющей степень радиационного воздействия, т.е. мерой ожидаемых последствий облучения объектов живой и неживой природы. Поглощенная доза характеризует не само излучение, а его воздействие на среду. Однако, для изучения влияния радиации на живые организмы эти
х единиц недостаточно, поскольку такое влияние зависит не только от плотности поглощенной энергии, но и от ее распределения в пространстве, точнее – от энергии, переданной частицами на единице длины их пробега. Для альфа-частиц, например, она в 20 раз выше, чем для гамма-квантов, и поэтому, при одинаковой поглощенной дозе облучение этими частицами примерно в 20 раз опаснее гамма-облучения. Чтобы учесть это, вводится понятие эквивалентной дозы, равной произведению поглощенной дозы на коэффициент каче
ства k, который характеризует действие данного вида радиации на живые организмы. Коэффициент качества показывает, во сколько раз ожидаемый биологический эффект больше, чем для излучения с ЛПЭ=3,5кэВ на 1мкм пути в воде. (ЛПЭ (линейная передача энергии) вдоль пути пробега ионизирующей частицы характеризует потерю энергии заряженных частиц на единицу пути вследствие ионизации и возбуждения.) Единица эквивалентной дозы в системе СИ – зиверт (Зв). Внесистемная единица: бэр – биологич
еский эквивалент рентгена; 1Зв=100бэр. Основные физические величины, используемые в радиационной биологии, их единицы: Рассмотрим физический смысл приведенных в таблице величин. 1. Экспозиционная доза. Отражает количество падающей на объект энергии излучения за время облучения. Вычисляется по формуле: где dQ – полный заряд ионов одного знака, возникающих в воздухе при полном торможении всех вторичных электронов, образованных фотонами в малом объеме воздуха; dM – масса воздуха в этом объеме. 2. Коэффициент п
оглощения излучения. Вычисляется по формуле: где dE – средняя энергия, переданная излучением веществу в некотором элементарном объеме, dm – масса вещества в этом объеме. 1 Гр = 100 рад. 3. Активность изотопа. 1 Беккерель соответствует 1 ядерному превращению в секунду. 4. Мощность поглощенной дозы. Используется для характеристики распределения поглощенной дозы во времени. Отражает количество энергии излучения, поглощаемое в единицу времени единицей массы вещества.
5. Эквивалентная доза. В какой-либо точке ткани определяется уравнением: Н = ДQN, где Д – поглощенная доза, Q и N – модифицирующие факторы. Q показывает, во сколько раз ожидаемый для данного вида излучения биологический эффект больше, чем действие рентгеновского излучения мощностью 250 кЭВ. Для гамма- и бета-излучения Q = 1, для альфа-излучения он равен 20. N – произведение всех других модифицирующих факторов. Т.е если внешнее излучение составляет 3 Р/ч, то человек находившийся
под этим воздействием получит за этот час суммарную дозу 3 бэра, если излучаются гамма- и бета-частицы, и 60 бэр, если излучаются альфа-частицы. Однако одинаковые количества поглощенной энергии дают часто разный биологический эффект в зависимости от вида ионизирующего излучения. Поэтому для оценки степени повреждающего действия ионизирующей радиации на биологические объекты пользуются коэффициентом относительной биологической эффективности - О
БЭ. Как видно из таблицы, повреждающее действие альфа-излучения, нейтронов и протонов в 10-20 раз больше, чем рентгеновских лучей, биологическое действие которых условно принято за 1. Коэффициенты относительной биологической эффективности - ОБЭ Рентгеновские и гамма-лучи 1 Бета-излучение 1 Альфа-излучение 10 n (нейтроны быстрые и медленные) 5-20 р (протоны) 10 Следует только помнить, что коэффициенты эти условны. Результат также зависит от выбора показателя, ко
торый берется для сравнения биологической эффективности. Например, ОБЭ можно устанавливать по проценту смертности, по степени гематогенных изменений, по стерилизующему действию на половые железы и т. д. Реакция организма на ионизирующее излучение зависит от величины экспозиционной дозы, выражаемой в рентгенах (Р) и поглощенной дозы, выражаемой в радах (рад), в единицах СИ (Гр). Зависимость повреждения от интенсивности общего облучения (Горизонтов П.
Д 1960) Примечание. Условия облучения: рентгеновские лучи, 180 кВ, 10 мА, фильтр 0,5 мм Сu и 1 мм А1; мощность дозы 13-60 Р/мин. Вид животного Минимальная смертельная доза, Р Доза половинной выживаемости, ЛД50 Абсолютная смертельная доза Мыши 200 350-400 550-800 Крысы 250-300 450-600 650-800 Морские свинки 200-300 40
0 Кролики 800 1100 1400 Кошки - - 550 Собаки 275 400 600 Обезьяны - - 600-700 Степень тяжести радиационного поражения зависит не только от дозы излучения, но и от длительности воздействия (мощности дозы). Повреждающее действие ионизирующей радиации при кратковременном облучении более выражено, чем при длительном облучении в одной и той же дозе. При дробном (фракционированном) облучении наблюдается снижение биологического эффекта: организм может переносить облучение в более высоких суммарных до
зах. Индивидуальная реактивность и возраст имеют также большое значение в определении тяжести радиационного поражения. В опытах на животных обнаружены широкие колебания индивидуальной чувствительности- одни собаки выживают после однократного облучения в дозе 600 Р, другие погибают после облучения в дозе 275 Р. Молодые и беременные животные более чувствительны к ионизирующему облучению. Старые животные также менее резистентны вследствие ослабления у них процес
сов регенераций. Влияние ИИ на иммунную систему Уникальная особенность ионизирующих излучений как этиологического фактора клинической патологии, состоит в том, что энергетически ничтожное в тепловом выражении (хотя и весьма значительное в единицах радиационной дозы) количество ионизирующей радиации, эквивалентное «энергии», заключенной в чашке горячего чая, поглощенное в едва уловимые доли секунды организмом человека или животного, может вызвать изменения, которые с неизбежностью реализуются в острую лучевую боле
знь, нередко со смертельным исходом. Указанный феномен, именуемый «энергетическим парадоксом», на заре радиобиологии называли «основным парадоксом радиобиологии». Его смысл долгое время оставался загадкой и начинает проявляться только сейчас. Становится понятным, как, через какие механизмы относительно малое на входе в организм количество энергии трансформируется во многообразные биологические и выраженные медицинские эффекты в зависимости от дозы. В основе этих эффектов лежат два критических события: 1) стойкие
Не поддающиеся устранению путем репарации, структурные повреждения генетического материала; 2) радиационно индуцируемые изменения в биомембранах, запускающие каскад стандартных ответных реакций клетки, направленных на поддержание генетической основы биологического вида. При этом особенно важно давнее соображение, реально подтверждающееся в последнее время: «Радиация не порождает каких-либо новых биологических феноменов; она лишь увеличивает вероятность возникновения различных клеточных событий, которые время от
времени происходят спонтанно». Как будут формироваться отдаленные эффекты облучения, возможны ли их прогнозирование и минимизация в группах повышенного риска – в значительной мере зависит от состояния иммунной системы. Ее можно охарактеризовать как полифункциональную, многоэтапно реализуемую систему по обеспечению надзора за осуществлением генетической программы и гомеостаза. Понятно, что иммунные механизмы принимают участие в развитии самых разнообразных патологических состояний у человека, выступая при э
том либо причиной, либо следствием. Индуцированные теми или иными воздействиями нарушения иммунитета ведут к дискоординации деятельности других регуляторных систем организма, что, в свою очередь, усугубляет несостоятельность системы иммунитета. Оценка последствий радиационного облучения для здоровья человека представляется крайне сложной проблемой, особенно это касается радиационных эффектов, возникающих при малых уровнях облучения. Результаты экспериментальных исследований, объективность которых обеспечивается
строго контролируемыми условиями проведения эксперимента, не всегда можно с достаточной надежностью экстраполировать на человека. Сложность этой проблемы обусловлена, помимо других, тремя обстоятельствами: 1) негомогенностью человеческой популяции с точки зрения индивидуальной радиочувствительности и ее изменчивости; 2) отсутствием единого взгляда ученых на реальный и гипотетический вред для здоровья человека низких уровней и интенсивности ионизирующего излучения; 3) отсутствием четких количественных характеристик этих у
ровней или диапазона так называемых малых доз ионизирующей радиации. Убедительные доказательства неоднородности и генетически детерминированной радиорезистентности (радиочувствительности) дают результаты иммуногенетических исследований, согласно которым существует тесная связь между воздействием ионизирующего излучения и риском реализации генетической предрасположенности к тем или иным патологическим состояниям. При изучении генетических систем крови участников ликвидации послед
ствий аварии на ЧАЭС обнаружены антигены, фенотипы и гаплотипы, которые ассоциируются с разной чувствительностью индивидуумов к радиационному воздействию. Крайние формы радиочувствительности у взрослых и детей могут различаться между собой многократно. В человеческой популяции 14-20% людей радиорезистентны, 10-20% – обладают повышенной радиочувствительностью и 7-10% – сверхрадиочувствительностью. К числу критических (высокочувствительных) органов по отношению к действию ионизирующей радиации принадлежит иммунная система. В о
стром периоде после облучения критичность иммунной системы определяется повреждающим воздействием на нуклеиновые кислоты, а также мембранные структуры иммунокомпетентных клеток за счет усиления перекисного окисления липидов, образования продуктов радиолиза воды и других активных соединений. Нарушение экспрессии дифференцировочных антигенов на мембранах клеток, принимающих участие в иммунном ответе, затрудняет их взаимодействие, ослабляет надзорную функцию иммунной системы. Существуют общие закономерности в измене
ниях качественного и количественного состава периферической крови под воздействием радиации. Снижение количества форменных элементов наступает тем раньше и интенсивней, чем больше доза облучения. Из-за высокой чувствительности клеток костного мозга, связанной с их интенсивным делением и дифференциацией, наблюдаются сильные изменения в периферической крови под воздействием радиации. Сравнительно небольшие дозы в 2 - 10 Гр вызывают гибель клеток костного мозга непосредственно в момент облучения или в митозах, при этом клетк
и теряют способность к делению. Генные перестройки в них в виде генных мутаций и хромосомных аберраций часто не мешают делению клетки. Элиминация мутантных клеток происходит медленнее, чем образование новых клеток, поэтому всегда имеется риск образования опухолей, особенно лейкозов. В костном мозге обнаруживаются следующие изменения: аплазия, фиброз, жировое его перерождение с островками кроветворной ткани, состоящей из зрелых гранулоцитов, через 6 месяцев после облучения обнаруживаются скопления ретикулярных кле
ток. Гипоплазия и аплазия костного мозга наблюдается в течение первых суток после облучения, что связано с массовой гибелью клеток. Нарушения выявляются сначала в гранулоцитопоэзе, затем в тромбоцитопоэзе, значительно позднее - в эритропоэзе. Наблюдается обеднение костного мозга ранними предшественниками кроветворения, т.к. эти клетки - малодифференцированные, интенсивно делящиеся, а следовательно и радиочувствительные. Поздние предшественники клеток периферической крови менее радиочувствительны, кроме предшествен
ников лейкоцитов и эритроцитов. Из-за резкого сокращения пула предшественников продукция зрелых форм в костном мозге временно снижается. Падение числа форменных элементов крови сопровождается включением компенсаторных механизмов, выражающихся в ускорении созревания клеток в костном мозге, уменьшении их жизнеспособности. Наблюдается относительное увеличение эритробластического роста. В ближайший период после лучевого воздействия наблюдается падение числа всех форменных элементов крови. Число циркулирующих эритроци
тов по данным одних авторов, уменьшается, другие исследователи приводят противоположные данные: в промежутке доз от 5 до 25 Р в крови крыс обнаруживается увеличение числа эритроцитов. Это явление увеличения показателей при облучении в малых дозах оправдано недавними исследованиями и получило название гормезиса. Предположительно, эффект усиления вызван стимуляцией центров нейро-эндокринной регуляции. Ряд исследователей отмечают снижение количества ретикулоцитов, что связано с укорочением их циркуляции и преобразованием
в зрелый эритроцит. Увеличения количества эритроцитов не происходит, так как значительно снижается продолжительность их жизни (до 43 дней). При визуальном исследовании мазков крови отмечалось снижение числа дискоцитов (нормальных эритроцитов) и увеличение содержания стоматоцитов, сфероцитов и шизоцитов. В целом количество аномальных форм эритроцитов через 5 лет после лучевой нагрузки достигало у ликвидаторов 25-30%. Эритроциты полихроматофильны, увеличивается их средний диаметр, средний объем и амплитуда анизоци
тоза. Снижается кислотная стойкость эритроцитов, чем объясняется снижение времени их циркуляции. Снижается способность костного мозга к синтезу гемоглобина. Со снижением числа эритроцитов закономерно падает и концентрация гемоглобина в периферической крови. Относительное содержание гемоглобина в одном эритроците увеличивается, увеличивается цветовой показатель. Изменяется количественный аминокислотный состав гемоглобина, ослабляется прочность связи между гемом и глобином, повышается процент метгемоглобина. Снижение
м количества гемоглобина после радиационного воздействия объясняется снижение кислородной емкости крови, при этом в 2-3 раза возрастает способность гемоглобина к включению соединений. Снижается содержание общего железа в плазме крови вследствие снижения числа эритроцитов. Увеличивается скорость включения железа в эритроциты и железо-связывающая способность плазмы. Снижается концентрация сывороточного ферритина, необходимого для синтеза гема. Регуляция эритропоэза осуществляется гормоном гликопротеиновой природы эр
итропоэтином. Он действует на клетки-предшественники эритроцитов, а также увеличивает скорость образования гемоглобина. Высокие дозы облучения вызывали обогащение крови эритропоэтинтормозящими веществами, хроническое облучение в малых дозах не вызывало каких-либо изменений в содержании эритропоэтинов. Увеличение количества СОЭ отмечено многими исследователями. Это может быть следствием снижения числа эритроцитов, снижения отрицательного заряда мембраны в сторону более положительного. При снижении количества ретикулоцитов СОЭ снижа
ется, т.к. ретикулоцит имеет более отрицательный поверхностный заряд, чем эритроцит. По видимому, в радиационном увеличении СОЭ основную роль играет снижение числа эритроцитов и изменение заряда их мембран. Костномозговое кроветворение (КМК) относится к тем системам, которые благодаря наличию стволового пула и относительной автономии пролиферации сравнительно быстро реагируют на воздействия ионизирующей радиации. При исследовании системы крови у животных после аварии на
ЧАЭС обнаружен ряд особенностей реакции КМК при постоянном пребывании их в условиях внешнего и внутреннего облучения в полях низкой интенсивности и различного качества. Наблюдения велись с июня 1986 г. на диких мигрирующих животных, на экспериментальных животных в условиях вивариев Чернобыля и Киева. Некоторые ветеринарные хозяйства организовали наблюдения над крупным рогатым скотом и овцами. Наблюдения продолжаются по настоящее время, особенно в местах с плотным загрязнением п
о цезию. Выявленные гематологические эффекты, как правило, превышают ожидаемые для дозовых нагрузок, исходящих из экстраполяции данных, описанных при воздействий на организм больших доз. Некоторые различия в степени выраженности изменений КМК обусловлены особенностями проведения опытов (временем, прошедшим после аварии, удаленностью от реактора места проведения экспериментов, продолжительностью самого эксперимента).
С октября 1986 г. систематически проводятся исследования КМК на белых беспородных крысах, завозимых в половозрелом трехмесячном возрасте в Чернобыль и наблюдаемых до момента их естественной гибели. Дозовые нагрузки не превышали 3 сГр за жизнь животного. К настоящему времени систематизирован материал о состоянии КМК у крыс трех серий экспериментов: 1986 - 1989, 1989 - 1991, 1991 - 1993 гг. Наиболее выраженные количественные изменения клеточного состава ко
стного мозга и периферической крови зарегистрированы в первой серии опытов. У животных чернобыльской группы отмечались: умеренно выраженная гипохромная анемия; прогрессирующая с третьего месяца пребывания в зоне лейкопения, преимущественно за счет лимфоцитарной фракции, достигающая к моменту гибели 30 - 40 % от исходного уровня; снижение числа миелокариоцитов на 50 - 60 %. Однако наиболее существенным было наличие гранулоцитопении с очень высоким содержанием эозинофилов. И
зменения в миелограмме наблюдались по гипопластическому типу (уменьшение молодых дифференцирующихся элементов с возрастанием доли зрелых гранулоцитов, ретикулярных и плазматических клеток). При этом в киевской группе животных гематологические изменения носили однонаправленный характер, но развивались значительно более медленно. В последующих сериях экспериментов ни в чернобыльской, ни в киевской группах на протяжении жизни животных не наблюдалось значительного снижения клеточности костного мозга и лейкоцитов
периферической крови. Обращает на себя внимание тот факт, что в каждой последующей серии снижается исходный уровень лейкоцитов. Это свидетельствует о тенденции к постоянному уменьшению плацдарма КМК под влиянием ухудшившейся радиоэкологической обстановки. К стабильным явлениям, наблюдаемым в каждой серии опытов, относятся относительная и абсолютная эозинофилия и наличие патологических клеток, обычных для радиационных поражений (гигантские гиперсегментированные нейтрофилы, клетки с фрагментозом ядер
Косматой структурой хроматина, включением ядерного вещества в цитоплазму, дву- и многоядерные лимфоциты, полиморфно-ядерные лимфоциты, мононуклеары и др.). Эозинофилию и атипичные клетки регистрируют практически все исследователи, изучающие систему КМК у животных. Наблюдаются они и у лиц, принимавших участие в ликвидации последствий аварии и у проживающих на загрязненных радионуклидами территориях. Этот феномен требует тщательного изучения, являясь показателем наличия в организме аутоиммунных реакций и развития эндогенной интоксикации. К ос
обенностям реакции КМК относятся также выявленные изменения пролиферативной активности костного мозга. У животных всех серий экспериментов после 3 - 6-месячного пребывания в Чернобыле отмечалось первичное значительное увеличение митотической активности, в ряде случаев сопровождаемое нарастанием клеточности костного мозга, с последующим выраженным снижением числа митозов. Механизм этого процесса остается невыясненным. Аналогичные результаты получены при изучении системы КМ
К у Диких грызунов, отлавливаемых в зоне аварии, получавших внешнее гамма-облучение от 5,16 10-9до 5,16 10-5Кл/кг. В реакции крови отмечено две фазы: усиление компенсаторных процессов (активация эритро- и миелопоэза) и декомпенсация (на фоне лейко- и эритропений происходит обильный выход в периферическое русло бластных форм и атипичных клеток). В работах приведены изменения гематологических показателей крупного рогатого скота, в течение 2 мес пребывав
шего на расстоянии 9 - 12 км от ЧАЭС. У животных наблюдалась эритропения, снижение концентрации гемоглобина, уменьшение процентного содержания нейгрофилов и моноцитов, эозинофилия, качественные изменения клеток белой крови. У крупного рогатого скота, свободно обитавшего в радиусе 3 - 6 км от аварийного блока до октября 1987 г в лейкограмме выявлена выраженная эозинофилия, сдвиг в формуле влево, лимфопения, наличие ретикулярных, недифференцированных клеток, фигур митозов, распадающихся форм; отмечалась гаперхромная анем
ия [б]. Количественные и качественные изменения показателей КМК отмечались и у лабораторных животных, кратковременно экспонированных в реперных точках 30-километровой зоны ЧАЭС. Например, отмечено, что у крыс после их экспонирования в течение 30 дней в с. Янов (доза 0,6 Гр) наблюдаются уменьшение лейкоцитов с 8,8 до 3,0 10-9 кл./л и тенденция к снижению клеточности костного мозга, содержание эритроцитов не изменилось.
У мышей, экспонируемых в этой же точке, в периферической крови обнаружили уменьшение числа лимфоцитов и лейкоцитов. Исследования по изучению стволового пула КМК единичны. По данным ряда авторов, у мышей, экспонируемых на реперных точках в 30-километровой зоне ЧАЭС в 1991 и 1992 гг. (суммарные дозы 24 и 120 мГр), наблюдается изменение стволовых потенций костного мозга. На основании экспериментов с дополнительным облучением животных в дозе
1,5 Гр установлено, что пребывание в зоне повышает радиочувствительность колониеобразующих единиц селезенки, т.е. отсутствует адаптивный ответ при остром облучении после экспозиции мышей в зоне аварии. Основные механизмы поражения КМК, по-видимому, следующие: 1) внешний гамма-фон всей совокупности среды обитания; 2) соприкосновение циркулирующей крови со всем спектром нуклидов, вдыхаемых легкими. При этом все газообразные и аэрозольные радионуклиды могут проникать в кровь через альвеолярные мембраны и мемб
раны сосудистого эндотелия традиционным путем транскапиллярного обмена. Иными словами, в сосудистом русле, в том числе и в межтрабекулярных полостях и синусах костного мозга, где располагаются кроветворные клетки, может создаваться определенная концентрация не тропных по отношению к костной или кроветворной ткани радионуклидов, постоянно воздействующих на организм, как бы Облучая "извне" кровь и кроветворные органы.
Тот же механизм попадания в кровь и соединений, содержащих радионуклиды, заглатываемые с пищей: через ворсинки кишечника, имеющие так назывемые окончатые капилляры, они попадают в систему воротной вены печени, откуда разносятся по всему организму, и, в свою очередь, являются одной из слагаемых лучевого воздействия на костномозговое кроветворение и периферическую кровь; 3) постоянное воздействие излучения на кровь и кроветворные органы имеет и геометрически "обратный" путь воздействия - либо непосредствен
но путем пробега частиц из фиксированных в тканях радиоизотопов, либо из радионуклидов, находящихся в тканях и клетках в растворимых соединениях, и проникающих обратно в кровь всеми классическими путями транскапиллярного обмена. Иными словами, осуществляется постоянный обмен энергии ионизирующего излучения между циркулирующей кровью и тканями организма с поддержанием относительного постоянства концентрации суммарных радионуклидов в капиллярах и синусоидах костного мозга, непосредственно кровоснабжающих межтрабекулярны
е полости, в которых содержатся стволовые и дифференцирующиеся элементы КМК; 4) действие остеотропных радионуклидов, таких, как90Sr и239Pu накапливающихся в эндостальной поверхности; кости, т.е. непосредственно прилегающей к трабекулярным поверхностям или поверхностям костномозговых каналов, при том, что все стволовые и бластные клетки костномозговой ткани располагаются строго по периферии. Кроме радиобиологических эффектов, развивающихся по канонам взаимодействия ионизирующего излучения от инкорпориро
ванного.источника с веществом живой ткани, 239Ри, обладая альфа-излучением, имеющим энергию, превышающую 5 мэВ с пробегом частиц до 250 мкм в жидкой фазе, будет обладать и выраженным прямым повреждающим действием на все клетки КМК с преобладанием повреждения стволового и коммитированных пулов, но могущим повредить клетки любой степени дифференцировки, в том числе и зрелые, а,также стромальные клетки кроветворного микроокружения; 5) и, наконец, соприкосновение всех классов дифференцирующихся в. костном мозге клеток строма
льного микроокружения, а также клеток периферической крови с "горячими" частицами, создающими.колоссальное энергетическое поле вокруг себя и обладающими очень большим прямым повреждающим действием, прямо зависящим от суммарной энергии ионизирующего излучения "горячей" частицы. Кроме перечисленных механизмов прямого повреждения клеток КМК инкорпорированными радионуклидами, в патогенезе костномозгового синдрома существенную роль играет развивающаяся эндогенная интоксикация.
Установлено, что радиационно-индуцированные мутации в локусе Т-клеточного рецептора (TСR) влияют на эффективность клеточного взаимодействия. Они могут быть использованы в качестве показателя биологической дозиметрии. В отдаленном периоде количество TCR-позитивных клеток прямо коррелирует со снижением иммунитета у пациентов, перенесших острую лучевую болезнь. Нарушение в отдаленном периоде после облучения иммунологических механизмов противоопухолевой резистентности, среди которых цитотоксичность естественных киллеров (ЕК) играет
ведущую роль, приводит к развитию стохастических онкологических эффектов. Результаты экспериментальных, клинических и эпидемиологических исследований свидетельствуют о высокой бластомогенной эффективности ионизирующей радиации. Рак возникает не сразу. Он является последним звеном длинной цепи изменений, которые нередко называют предраковыми или предопухолевыми заболеваниями. Обнаружены некоторые особенности взаимодействия клеток стромы и гемопоэтических клеток костного мозга, обусловленные воздействием ионизирующего излучения.
В частности, отмечаются блокирование лимфоцитов в стромальных элементах, а также активация процесса разрушения мегакариоцитов нейтрофильными гранулоцитами. Не исключено, что длительно сохраняющиеся структурные и функциональные изменения в клетках стромы под воздействием ионизирующей радиации инициируют злокачественную трансформацию. Вопрос о роли стромы в развитии в отдаленный период после облучения гематологической патологии, в частности миелодиспластического синдро
ма и лейкемии, в силу его особой важности требует дальнейшего изучения. Несмотря на высокий регенерационный потенциал большинства клеточных компонентов иммунной системы, восстановление затягивается на годы, особенно у реконвалесцентов острой лучевой болезни. Причем изменения не всегда имеют четкую зависимость от дозы облучения, которую в классической радиобиологии считали и продолжают считать единственно верным доказательством ответа биологической системы на воздействие ионизирующей радиации. Иммунодефицит, как
конечная или существенно продвинутая во времени патогенетическая стадия изменений в иммунной системе пострадавших вследствие радиационной аварии, определяется достаточно редко. Чаще выявляют в разной степени выраженную количественную или функциональную недостаточность тех или иных субпопуляций клеток или нарушение продукции гуморальных факторов с реализацией на уровне организма в виде соматической патологии – заболеваний пищеварительной, нервной, сердечно-сосудистой, дыхательной и выделительной систем. Отмеча
ют значительное увеличение частоты выявления аллергических заболеваний (до 20%) и клинических проявлений иммунной недостаточности (до 80%) у лиц, облученных в дозе свыше 0,25 Гр. Одним из приоритетных вопросов, требующих неотложной научной разработки, являются персистирующие вирусные инфекции у пострадавших контингентов. Результаты обследования больных со стойкими лимфоцитозом и лейкопенией, связанными с влиянием облучения, в 2/3 случаев выявляли наличие персистирующих инфекций, цитомегаловирусной, токсоплазменной и др что д
ало возможность провести адекватное лечение и иммунологическую коррекцию. Следует заметить, что подходы к иммунокоррекции должны быть строго индивидуализированы, обоснованы соответствующим объемом исследований, так как первоначальные выводы о радиационно-обусловленных нарушениях иммунной системы, наличии иммунодефицитного состояния и необходимости иммуностимулирующей терапии, сделанные в медицинских учреждениях городского или районного уровня на основании наблюдения за пациентами, после экспертной оценк
и подтверждались только у 15,2% больных. Человеческий организм – единое целое, в условиях аварии и послеаварийных событий раннего и отдаленного периода он подвергается, помимо радиации, воздействию других факторов нерадиационной природы. Психогенный стресс – один из наиболее мощных в этом ряду. Выявлено, что воздействие стресса на нейроэндокринную систему сопровождается увеличением в крови нейропептидов, катехоламинов, глюкокортикоидов и других гормонов гипоталамо-гипофизарно-надпочечниковой оси.
Высокий уровень в крови глюкокортикоидов и других гормонов вызывает инволюцию тимуса, уменьшение количества лимфоцитов селезенки, костного мозга, снижение активности макрофагов, пролиферации лимфоцитов и повышение продукции цитокинов. Однако не только нейроэндокринная система влияет на функции иммунной системы, но и, наоборот, иммунная система воздействует на гипоталамо-гипофизарно-надпочечниковую ось через рецепторы для цитокинов. К факторам нерадиационной природы относятся также промышленные и бытовые алле
ргены, соли тяжелых металлов, компоненты выхлопных газов транспорта и др. Следовательно, мы вправе говорить о комплексном экологически неблагоприятном воздействии на организм, отражающимся на деятельности иммунной системы. Данные исследований тиреоидной системы пострадавших в остром так называемом «йодном периоде» аварии выявили изменения, характерные для постепенного развития нестохастических эффектов облучения щитовидной железы. Иммунные сдвиги в период первичной тиреоидной реакции свидетельствовали о начале
развития хронических, с большей вероятностью аутоиммунных, тиреоидитов. Группу повышенного риска развития хронического тиреоидита и гипотиреоза составили пациенты, перенесшие облучения щитовидной железы наиболее сложного комбинированного характера: сочетание внутреннего облучения короткоживущими изотопами йода с внешним γ-облучением. Эту группу составили бывшие жители 30-километровой зоны ЧАЭС и участники ликвидации последствий аварии «йодного периода» 1
986 г. В клинико-экспериментальных исследованиях установлено, что развитие нейроаутоиммунных реакций может быть одним из звеньев патогенеза послерадиационной энцефалопатии. Неоднозначны оценки медицинских последствий для здоровья пострадавшего населения от атомных бомбардировок японских городов Хиросимы и Нагасаки. Однако в последние годы приводят доказательства значительного ухудшения состояния здоровья «хибакуши» по сравнению со стандартной японской популяцией по многим классам болезней (в 1,7-13,4 раза). По м
нению исследователей, увеличение распространенности заболеваний, включая рак и лейкемию, реализация которых обусловлена сбоями в полифункциональной деятельности иммунной системы, связано с воздействием ионизирующего излучения в те годы, когда эти больные были детьми или молодыми людьми. Исследования иммунного статуса детей и подростков, пострадавших вследствие чернобыльской катастрофы, занимают особое место в общей проблеме пострадиационных эффектов. Осуществленный в рамках национальной программы «Дети Чернобыля»
многолетний мониторинг состояния иммунной системы у лиц, облученных в детском возрасте в результате воздействия радионуклидов йода (131І, 129І), а также 137Cs, 90Sr, 229Pu и др позволил установить определенные закономерности в этапности развития дозозависимых изменений в иммунной системе и функции щитовидной железы. Результаты выполненных в первые послеаварийные годы исследований иммунной системы у детей, проживающих на загрязненных радионуклидами территориях, свидетельствуют
о наличии нерезко выраженных, но статистически достоверных отклонений в субпопуляциях Т- и В-лимфоцитов от соответствующих показателей контрольной группы пациентов. На этапе наблюдений в 1991-1996 гг. были выявлены различия между группами облученных и необлученных детей по уровню содержания главных регуляторных субпопуляций лимфоцитов периферической крови и направленности корреляционной зависимостью между содержанием Т В-клеток, ЕК, CD3+, CD4+ Т-клеток и дозами облучения щитовидной железы радиоиодом
Начиная с 1994-1996 гг были получены убедительные данные о развитии 131І-дозозависимых аутоиммунных нарушений, основанные на фенотипической оценке лимфоцитов по главным локусам гистосовместимости HLA, HLA-Dr и многим другим параметрам лимфоцитарных субпопуляций. Ретроспективный анализ состояния иммунной системы детей, проживающих на загрязненных радионуклидами территориях, свидетельствует о манифестации иммунодефицитных нарушений преимущественно по смешанному типу. Установлено, что 68% детей с отклонениями в
иммунном статусе имеют генетические аллели, контролирующие направленность иммунного реагирования организма, и которые, как правило, связаны с низким ответом иммунной системы на действие любых экзогенных факторов, либо с аутоиммунными процессами. Это, прежде всего, антигены HLA-A9, HLA-B7, HLA-DR4, HLA-Bw35, HLA-DR3, HLA-B8. На основании полученных результатов можно полагать, что у этих детей реализовалась генетическая предрасположенность к иммунным нарушениям вследствие воздействия
экологически неблагоприятных факторов, в частности радиационного. По сравнению со взрослыми, в формировании тиреоидных нарушений у детей преобладающая роль принадлежит антигену HLA-Bw35, который одновременно является маркером аутоиммунных процессов. Следует также отметить, что степень ассоциативной взаимосвязи антигенов гистосовместимости с заболеваниями в детском возрасте значительно выше, чем у взрослых. Результаты иммуногенетических и иммуноцитологических исследований подтверждены клиническими проявлениями радиационно-индуциров
анных нарушений функции щитовидной железы, а также данными эпидемиологических исследований, проведенных у более 10 тыс. детей, облученных в «йодном периоде» (эвакуированных из 30-километровой зоны аварии) и свыше 2,5 тыс. детей – жителей радиоактивно загрязненных территорий (облученных в «йодном периоде» и постоянно подвергающихся облучению за счет долгоживущих радионуклидов 137Cs, 90Sr и др. Получены данные о негативном влиянии малых доз ионизирующей радиации на противодифтерийный,
противостолбнячный, противокоревой и противококлюшный иммунитет у детей, проживающих на загрязненных радионуклидами территориях. Это обосновывает создание дифференцированных программ иммунизации с учетом региональных и индивидуальных особенностей иммунного статуса детей. Исследования, проведенные после 2001 г указывают на дозозависимые эффекты в иммунной системе даже через 15 лет, а порог воздействия ионизирующей радиации на иммунную систему по большинству изученных параметров сост
авляет 250 мЗв. При оценке функциональной активности лимфоцитов крови и периферических лимфоидных органов выявлены: нарушения реакции на поликлональный Т-клеточный митоген при одновременной активации функции К-клеток (антителозависимая цитотоксичность); подавление кооперативных Т-клеточных реакций - трансплантационного иммунитета, гиперчувствительности замедленного типа. Достаточно типичны волнообразные изменения способности лимфоцитов к контактному взаимодействию с аллогенными тканевыми базофилами. Такое взаимодейств
ие, согласно современным представлениям, определяется степенью дифференцировки лимфоидных клеток и опосредует их участие в регуляции аллергических реакций немедленного и замедленного типов, а также регуляции гуморального иммунитета. К биологическим эффектам постоянного облучения относится и прогрессирующее со временем снижение "спонтанной" антигеннеспецифической Т-супрессии. Показатели, характеризующие В-систему иммунитета, более стабильны. При обследовании нескольких поколений линейных мышей, постоя
нно содержащихся в Чернобыле, не выявлено существенных изменений содержания и пролиферативной активности В-лимфоцитов в периферических лимфатических узлах. Ответ на поликлональный В-митоген (декстрансульфат) и уровни сывороточных иммуноглобулинов, а также специфический гуморальный иммунный ответ на заражение вирусом гриппа у этих животных также значительно не изменялись. О сохранении способности к активному антителообразованию свидетельствует и выраженная стимул
яция аллергической реакции немедленного типа - достоверное повышение содержания Іg E-антител в органах дыхания мышей в ответ на иммунизацию аллергеном амброзии. В отдаленные сроки радиационного воздействия выявлено и возрастание уровня аутоантител к собственным эритроцитам и эпителиальному ретикулуму тимуса. Эти данные указывают не только на большую сохранность гуморального иммунитета по сравнению с клеточным, но и на срыв толерантности к собственным тканям. Последнее свид
етельствует о высокой вероятности развития аутоиммунных поражений облученного организма. Поздние реакции на постоянное действие факторов радиационной аварии другой группы клеток иммунной системы - моноцитов (макрофагов) - изучены в меньшей степени. Известно, что в костном мозгу страдают клетки моноцитарного ряда кроветворения. Выявлены усиление поглотительной активности макрофагов брюшной полости и активация ферментов "дыхательного взрыва" фагоцитирующих клеток до критических уровней. Нар
яду с этим обнаруживается четкая тенденция к истощению функционального резерва клеток. В детальном изучении на экспериментальных моделях нуждается продукция моноцитами (макрофагами) цитокинов, играющих важную роль в развитии воспалительной реакции, в процессах пролиферации и дифференцировки клеток иммунной системы, в противоопухолевой резистентности, межсистемных взаимодействиях, развитии компенсаторных процессов. Важность фундаментальных исследований такого рода, помимо теоретических посылок, обусловлена и тем,
что к настоящему времени показано, что у ликвидаторов к числу отдаленных эффектов относится изменение концентрации сывороточных цитокинов этой группы (в первую очередь ИЛ-1b). Получены предварительные результаты об изменении уровней регуляторных цитокинов и у постоянно облучаемых экспериментальных животных. При изучении показателей естественной резистентности выявлено снижение лизоцимной активности сыворотки крупного рогатого скота.
При интегральной оценке иммунологической реактивности животных, постоянно подвергающихся действию вредных факторов аварии на ЧАЭС, установлено развитие иммунодефицитов, проявлениями которых являются: снижение антимикробной устойчивости кожи у крупного рогатого скота и диких мышевидных грызунов; повышение чувствительности к экспериментальным вирусным инфекциям и прививке экспериментальных штаммов опухолевых клеток у лабораторных мышей. Важно отметить, что развитие иммунодефицитных состояний наблюдается на протя
жении всего срока проведения исследований (1986- 1993), т.е. иммунодепрессивное действие факторов аварии на ЧАЭС - долговременный биологический эффект. Существенное теоретическое и практическое значение имеет тот факт, что, хотя иммунологическая недостаточность развивается вне зависимости от того, в каком периоде индивидуального развития началось радиационное воздействие, степень нарушений иммунного статуса возрастает,
а сроки их появления сокращаются тем значительнее, чем моложе организм. Наибольшие изменения иммунной реактивности обнаружены у потомков облученных родителей, в свою очередь начинающих подвергаться постоянному облучению уже с эмбрионального периода. Анализ динамики изменений в иммунной системе показывает, что на ранних этапах (первые месяцы) постоянного облучения в малых дозах наряду с повреждающим действием факторов радиационной аварии наблюдаются признаки функциональной напряженности, компенсаторной и репар
ативной реакций. За счет последних отдельные иммунологические показатели могут превышать контрольный уровень, создавая, на первый взгляд, впечатление активации иммунной системы. Однако полноценная адаптация системы иммунитета, видимо, отсутствует, ее компенсаторно-репаративные возможности истощаются и, по мере увеличения возраста животных либо числа поколений, выявляются нарушения преимущественно деструктивного характера, а также значительные нарушения иммунного гомеостаза.
Изучение возрастной динамики характеристик иммунокомпетентных органов и иммунных реакций у животных, содержащихся на экспериментальной базе в Чернобыле, позволило высказать предположение об ускорении темпов старения иммунной системы при постоянном внешнем и внутреннем облучении малыми дозами радиации низкой интенсивности. Ускоренное развитие возрастной супрессии прямо показано в модельных экспериментах на линейных мышах, облучаемых дважды в неделю дозой 0,07 Гр в течение месяцев. Следует отметить, что признаки "рад
иационного старения" тимуса обнаружены при обследовании облученных - "ликвидаторов" и пациентов с проявлением отдаленных последствий острой лучевой болезни. Совокупность результатов, полученных разными исследованиями, показывает, что структура и выраженность иммунодефицитов может очевидно, варьировать в зависимости от дозы облучения, спектра и распределения радионуклидов в организме, генетических особенностей (данные, полученные на инбредных мышах разных линий) и исходного физиологического состояния последнего. Однако в б
ольшинстве исследований наблюдается преимущественная связь отдаленных эффектов факторов Чернобыльской катастрофы с повреждением тимусзависимого (Т-) звена системы иммунитета. Важно, что подобные закономерности выявлены и в исследованиях, посвященных иммунному статусу взрослого и детского населения, пострадавшего в результате аварии на ЧАЭС, в том числе лиц, принимавших участие в ликвидации последствий аварии. На основании накопленной в настоящее время информации можно предполагать, что патоген
ез постчернобыльских иммунодефицитов имеет, вероятно, сложный комплексный характер и включает ряд компонентов: прямое и непрямое повреждающее действие ионизирующих излучений на клетки иммунной системы, включая стромальные элементы и вспомогательные клетки; нарушение процессов созревания и дифференцировки иммунокомпетентных клеток (преимущественно Т-лимфоцитов); нарушение функций центрального органа иммунитета - тимуса; развитие аутосенсибилизации (в том числе и к клеткам эпителиального ретикулума тимуса); глубокую
дискоординацию иммунорегуляторных процессов и взаимодействий в рамках иммунной системы; изменение гормональной регуляции иммунной реактивности, связанное с нарушением в эндокринной системе. Приведенный перечень может быть далеко не исчерпывающим; не вполне ясно также, какие из перечисленных феноменов первичны, а какие вторичны. Однако более детальное обсуждение механизмов развития иммунодефицитов при постоянном воздействии на организм млекопитающих факторов аварии на ЧАЭС преждевременно.
Выводы Изменениям, которые происходят в иммунной системе, занимающей промежуточное место между критическими и некритическими системами организма, принадлежит особая роль в патогенезе лучевой болезни. Наиболее демонстративным проявлением радиационного повреждения иммунитета признаны иммунодефицит и повышение чувствительности к возбудителям инфекционных заболеваний, сопровождающиеся количественными и качественными изменениями нормальной микрофлоры организма, в частности кишечника.
Причинами иммунодепрессии и иммунодефицита, развивающихся вскоре после облучения, является гибель, повреждение функции и миграционных свойств лимфоцитов, а также нарушение количественного соотношения субпопуляций лимфоцитов, а также нарушение количественного соотношения субпопуляций лимфоцитов и их функциональных взаимодействий. Нарушение нормальных количественных соотношений субпопуляций лимфоцитов обусловлено их различной чувствительностью: В- клетки более радиочувствительные, чем Т-клетки; однако числ
енность В-клеток восстанавливается быстрее, чем численность Т-клеток. Нарушение антимикробного иммунитета и связанные с ним инфекционные осложнения, кроме того, можно рассматривать как следствие повышения проницаемости тканевых барьеров, нарушения фагоцитарной способности клеток ретикулоэндотелиальной системы и угнетения неспецифических бактерицидных систем организма – пропердина, лизоцима, бактерицидных субстанций ряда тканей, а также бактерицидности кожи. Кроме того, облучение угнетает образование новых антител. Большое значение имеют такж
е развивающиеся в облученном организме аутоиммунные процессы, составляющие самостоятельную проблему неинфекционной иммунологии. Аутоантигенами в принципе могут быть как нормальные ткани при их попадании в русло крови, где они обычно не встречаются, так и патологически измененные белки и связанные с ними вещества. Полсе облячения создается реальная возможность столкновения организма с аутоантигенами обоих видов вследствие быстро развивающейся тканевой деструкции, резкого повышения проницаемости биологических барьеров и измен
ения антигенных свойств тканей. Несомненно, интеграция знаний в области иммунологии и радиобиологии, произошедшая в результате ядерной катастрофы, явилась своеобразным стимулом в формировании и развитии нового научно-клинического направления – радиационной иммунологии. Масштабность и многогранность медицинских последствий чернобыльской катастрофы катализировали многочисленные экспериментальные и клинические исследования, что способствовало не только накоплению фактов,
но и обеспечило получение значимых научных выводов и практических рекомендаций для клинической иммунологии. Сегодня представляется очевидным спад интереса мировой общественности к проблемам, связанным с чернобыльской аварией. Это обусловлено появлением новых серьезных гуманитарных проблем, требующих безотлагательного решения. Вместе с тем, атомная энергетика продолжает развиваться, что обусловлено все возрастающими потребностями человечества в энергоресурсах, и, соответственно, постоянно увеличивается количество лю
дей, имеющих профессиональные контакты с ионизирующим излучением. К концу прошлого века в развитых странах их число приблизилось к 7-8% населения. Поэтому проблема влияния ионизирующей радиации на иммунную систему человека и в перспективе будет иметь большое практическое значение. Список литературы 1. Антипкин Ю.Г Чернышов В.П Выхованец Е.В. Радиация и клеточный иммунитет у детей
Украины. Обобщение данных І и начала ІІ этапов десятилетнего (1991-2001 гг.) мониторинга состояния иммунной системы у детей и подростков, пострадавших от облучения вследствие аварии на Чернобыльской АЭС // Международный журнал радиационной медицины. – 2001. – № 3-4. – С. 152. 2. Бебешко В.Г Базика Д.А Кліменко В.І. та ін. Гематологічні та імунологічні ефекти хронічного опромінення // Чорнобиль: Зона відчуження / Під ред.
В.Г. Бар"яхтара. – К.: Наукова думка. – 2001. – C. 214-216. 3. Верещагина А.О Замулаева И.А Орлова Н.В. и др. Частота лимфоцитов, мутантных по генам Т-клеточного рецептора как возможный критерий для формирования групп повышенного риска развития опухолей щитовидной железы у облученных и необлученных лиц // Радиационная биология, радиоэкология. – 2005. – Т. 45. – № 5. – С. 581-
586. 4. Минченко Ж.Н Базыка Д.А Бебешко В.Г. и др. HLA-фенотипическая характеристика и субпопуляционная организация иммунокомпетентных клеток в формировании пострадиационных эффектов в детском возрасте // Медицинские последствия аварии на Чернобыльской атомной станции. Монография в 3 книгах. Клинические аспекты Чернобыльской катастрофы. Книга 2. – К.: «Медэкол» МН
ИЦ БИО-ЭКОС. – 1999. – С. 54-69. 5. Орадовская И.В Лейко И.А Оприщенко М.А. Анализ состояния здоровья и иммунного статуса лиц, принимавших участие в ликвидации последствий аварии на Чернобыльской АЭС // Международный журнал радиационной медицины. – 2001. – № 3-4. – С. 257. 6. Потапова С.М Кузьменок О.И Потапнев М.П Смольникова В.В. Оценка состояния Т-клеточного и моноцитарного звеньев у ликвидаторов аварии на Чернобыльской АЭС чер
ез 11 лет // Иммунология. – 1999. – № 3. – С. 59-62. 7. Талько В.В. Показатели клеточного иммунитета, неспецифической резистентности и метаболическая характеристика иммунокомпетентных клеток при аутоиммунном тиреоидите у облученных в связи с аварией на Чернобыльской АЭС // Проблемы радиационной медицины. Респ. межвед. сб. – К. – 1993. – Вып. 5. – С. 41-45. 8. Чумак А.А. Иммунная система пострадавших «чернобыльцев» в отдале
нный послеаварийный период – диагностика недостаточности и подходы к коррекции // Международный журнал радиационной медицины. – 2001. – № 3-4. – С. 400. 9. Чумак А.А Базыка Д.А Коваленко А.Н. и др. // Иммунологические эффекты у реконвалесцентов острой лучевой болезни – результаты тринадцатилетнего мониторинга / Международный медицинский журнал. – 2002. – № 1 (5). – С. 40-41. 10. Ярмоненко С.П. Радиобиология человека и животных:
Учеб. Для биол. спец. вузов. – 3-е изд перераб. и доп М.: Высш. шк 1988. – 424 с.
Влияние радиации на иммунную систему и их последствия
Ионизирующие излучение в любых дозах вызывает функциональные и морфологические изменения в клеточных структурах и изменяет деятельность почти во всех системах организма. В результате этого повышается или угнетается иммунологическая реактивность животных. Иммунная система является высокоспециализированной, ее составляют лимфоидные органы, их клетки, макрофаги, клетки крови (нейтрофильные, эозинофильные и базофильные, гранулоциты), система комплемента, интерферон, лизоцим, пропердин и другие факторы. Главным иммунокомпетентными клетками являются Т - и В-лимфоциты, ответственные за клеточный и гуморальный иммунитет.
Направленность и степень изменений иммунологической реактивности животных при действии радиации определяется главным образом поглощенной дозой и мощностью облучений. Малые дозы излучения повышают специфическую и неспецифическую, клеточную и гуморальную, общую и иммунобиологическую реактивность организма, способствуют благоприятному течению патологического процесса, повышают продуктивность скота и птиц.
Ионизирующие излучение в сублетальных и летальных дозах приводит к ослаблению животных или угнетению иммунологической реактивности животных. Нарушение показателей иммунологической реактивности отмечается значительно раньше, чем проявляются клинические признаки лучевой болезни. С развитием острой лучевой болезни иммунологические свойства организма все более ослабляются.
Понижается резистентность облеченного организма к возбудителям инфекции может по следующим причинам: нарушение проницаемости мембран тканевых барьеров, снижение бактерицидных свойств крови, лимфы и тканей, подавление кроветворения, лейкопения, анемия и тромбоцитопения, ослабление фагоцитарного механизма клеточной защиты, воспаления, угнетения продукции антител и другие патологические изменения в тканях и органах.
При воздействии ионизирующего излучения в небольших дозах изменяется проницаемость тканей, а при сублетальной дозе и более резко увеличивается проницаемость сосудистой стенки, особенно капилляров. После облучения среднелетальными дозами у животных развивается повышенная проницаемость кишечного барьера, что является одной из причин расселения кишечной микрофлоры по органам. Как при внешнем, так и при внутреннем облучении отмечается увеличения аутофлоры кожи, которое проявляется рано, уже в латентный период лучевого поражения. Этот феномен прослеживается у млекопитающих, птиц и человека. Усиленное размножение и расселение микроорганизмов на коже, слизистых оболочках и в органах обуславливается снижением бактерицидных свойств жидкостей и тканей.
Определение числа кишечных палочек и особенно гемолитических форм микробов на поверхности кожи и слизистых оболочках является одним из тестов, позволяющих рано установить степень нарушения иммунобиологической реактивности. Обычно повышение аутофлоры происходит синхронно с развитием лейкопении.
Закономерность изменений аутофлоры кожи и слизистых оболочек при внешнем облучении и инкорпорации различных радиоактивных изотопов сохраняется. При общем облучении внешними источниками радиации наблюдается зональность нарушения бактерицидных кожных покровов. Последнее, по-видимому, связано с анатомофизиологическими особенностями различных участков кожи. В целом бактерицидная функция кожи находится в прямой зависимости от поглощенной дозы излучения; при летальных дозах она резко снижается. У крупного рогатого скота и овец, облеченных гамма-лучами (цезий-137) в дозе ЛД 80-90/30 , изменения аутофлоры кожи и слизистых оболочек начинается с первых суток, а к исходному состоянию у выживших животных приходят на 45-60-му дню.
Внутреннее облучение, как и внешнее, вызывает значительное понижение бактерицидности кожи и слизистых оболочек при однократном введении йода-131 курам в дозах 3 и 25 мКи на 1 кг их массы количество бактерий на коже начинает уже с первых суток увеличиваться, достигая максимума на пятый день. Дробное веление указанного количество изотопа в течении 10 дней приводит к значительно большому бактериальному обсеменению кожи и слизистой оболочки ротовой полости с максимумом на 10-й день, причем в основном возрастает число микробов с повышенной биохимической активностью. В следующее время прослеживается прямая связь численного увеличения бактерий с клиническим проявлением лучевого поражения.
Одним из факторов, обеспечивающих естественную антимикробную устойчивость тканей, является лизоцим. При лучевом поражении содержание лизоцима в тканях и крови уменьшается, что свидетельствует об уменьшении его продукции. Этот тест может быть использован для определения ранних изменений резистентности облеченных животных.
Большую роль в невосприимчивости животных к инфекциям играет фагоцитоз. При внутреннем и внешнем облучениях в принципе изменения фагоцитарной реакции имеют аналогичную картину. Степень нарушения реакции зависит от величины дозы воздействия; при малых дозах (до 10-25 рад) отмечается кратковременная активация фагоцитарной способности фагоцитов, при полулетальных - фаза активации фагоцитов сокращается до 1-2 дней, в дальнейшем активность фагоцитоза понижается и в летальных случаях доходит до нуля. У выздоравливающих животных происходит медленная активация реакции фагоцитоза.
Значительные изменения в облученном организме претерпевают фагоцитарные способности клеток ретикулоэндотелиальной системы и макрофагов. Эти клетки довольно радиорезистентны. Однако фагоцитирующая способность макрофагов при облучении нарушается рано. Угнетение фагоцитарной реакции проявляется незавершенностью фагоцитоза. По-видимому, облучение нарушает связь между процессами захвата частиц макрофагами и ферментативными процессами. Подавление функции фагоцитоза в этих случаях может быть связано с угнетением выработки соответствующих опсонинов лимфойдной системой, ибо известно, что при лучевой болезни отмечается уменьшение в крови комплемента, пропердина, опсонинов и других биологических веществ.
В иммунологических механизмах самозащиты организма большую роль играют аутоантитела. При радиационных поражениях происходит повышение образования и накопления аутоантител. После облучения в организме можно обнаружить иммунокомпетентные клетки с хромосомными транслокациями. В генетическом отношении они отличаются от нормальных клеток организма, т.е. являются мутантами. Организмы, в которых существуют генетически различные клетки и ткани, обозначаются как химеры. Образовавшиеся под действием облучения аномальные клетки, ответственные за иммунологические реакции, приобретают способность вырабатывать антитела против нормальных антигенов организма. Иммунологическая реакция аномальных клеток против собственного организма может вызвать спленомегалию с атрофией лимфоидного аппарата, анемию, отставание в росте и массе животного и ряд других нарушений. При достаточно большом количестве таких клеток может произойти гибель животного.
Согласно иммуногенетической концепции, выдвинутой иммунологом Р.В. Петровым, наблюдается следующая последовательность процессов лучевого поражения: мутагенное действие радиации→относительное увеличение аномальных клеток, обладающих способностью к агрессии против нормальных антигенов→накопление таких клеток в организме→аутогенная агрессия аномальных клеток против нормальных тканей. По мнению некоторых исследователей, рано проявляющиеся в облученном организме аутоантитела участвуют в повышении его радиорезистентности при однократных воздействиях сублетальных доз и при хроническом облучении малыми дозами.
О нарушении резистентности у животных при облучении свидетельствуют лейкопения и анемия, подавление деятельности костного мозга и элементов лимфоидной ткани. Поражение клеток крови и других тканей и изменение их деятельности сказываются на состоянии гуморальных систем иммунитета - плазме, фракционном составе сывороточных белков, лимфе и других жидкостях. В свою очередь, эти субстанции, подвергаясь воздействию излучения, оказывают влияние на клетки и ткани и сами по себе обуславливают и дополняют другие факторы снижения естественной резистентности.
Угнетение не специфического иммунитета у облученных животных приводит к усилению развития эндогенной инфекции - увеличивается количество микробов аутофлоры кишечника, кожи и других областей, изменяется ее видовой состав, т.е. развивается дисбактериоз. В крови и внутренних органах животных начинают обнаруживаться микробы - обитатели кишечного тракта.
Бактериемия имеет исключительно важное значение в патогенезе лучевой болезни. Между началом возникновения бактериемии и сроком гибели животных наблюдается прямая зависимость.
При радиационных поражениях организма изменяется его естественная устойчивость к экзогенным инфекциям: туберкулезным и дизентерийным микробам, пневмококкам, стрептококкам, возбудителям паратифозных инфекций, лептоспироза, туляремии, трихофитии, кандидамикоза, вирусам инфлюэнцы, гриппа, бешенства, полиомиелита, ньюкаслской болезни (высококонтагиозная вирусная болезнь птиц из отряда куриных, характеризующаяся поражением органов дыхания, пищеварения и центральной нервной системы), простейшими (кокцидиями), бактериальным токсинам. Однако видовая невосприимчивость животных к инфекционным болезням сохраняется.
Лучевое воздействие в сублетальных и летальных дозах отягощает течение инфекционной болезни, а инфекция, в свою очередь, утяжеляет течение лучевой болезни. При таких вариантах симптомы болезни зависят от дозового, вирулентного и временного сочетания действия факторов. При дозах облучения, вызывающих тяжелую и крайне тяжелую степень лучевой болезни, и при инфицировании животных первые три периода ее развития (период первичных реакций, латентный период и разгар болезни) в основном будут преобладать признаки острого лучевого заболевания. Заражение животных возбудителем остропротекающей инфекционной болезни незадолго или на фоне облучения сублетальными дозами приводят к утяжелению течения данной болезни с развитием относительно характерных для нее клинических признаков. Так, у поросят, облученных смертельными дозами (700 и 900 Р) и зараженных через 5 ч, 1,2,3,4, и 5 сут. после облучения вирусом чумы, при вскрытии находят в основном изменения, которые наблюдаются у облученных животных. Лейкоцитарная инфильтрация, клеточно-пролиферативная реакция, инфаркты селезенки, наблюдаемые при чистой форме чумы, в этих случаях отсутствуют. Повышенная чувствительность подсвинков к возбудителю рожи у переболевших лучевой болезнью средней степени тяжести сохраняется спустя 2 мес. после облучения рентгеновскими лучами в дозе 500 Р. При экспериментальном заражении возбудителем рожи болезнь у свиней проявляется более бурно, генерализация инфекционного процесса наступает на третьи сутки, тогда как у контрольных животных она обычно регистрируется только на четвертый день. Патоморфологические изменения у облученных животных при этом характеризуются выраженным геморрагическим диатезом.
Экспериментальными исследованиями на морских свинках и овцах выявлено своеобразное течение сибирской язвы у животных, больных лучевой болезнью средней тяжести. Как внешне, так и сочетанное воздействие радиации снижает у них устойчивость к заражению возбудителем данной болезни. Клинические признаки не являются строго специфичными ни для лучевой болезни, ни для сибирской язвы. У больных отмечается выраженная лейкопения, повышается температура тела, учащается пульс и дыхание, нарушается функция желудочно-кишечного тракта, в сыворотке крови проявляются сибиреязвенные антитела в низких титрах, выявляемые реакцией непрямой гемагглютинации. Болезнь протекает остро и заканчивается летально. При патологическом вскрытии во всех случаях регистрируется уменьшение селезенки и обсеменение сибиреязвенными микробами внутренних органов и лимфоузлов.
Следовательно, воздействие ионизирующего излучения на животных в сублетальных в летальных дозах вызывает снижение всех естественных факторов устойчивости организма к эндогенным и экзогенным инфекциям. Это проявляется тем, что у облученных животных возникновение инфекционных болезней происходит при меньшей дозе возбудителя, среди облученных увеличивается процент заболевающих, болезнь быстрее и чаще заканчивается гибелью.
Нарушение иммунобиологической реактивности возникают уже в период первичных реакций на облучение и, постепенно увеличиваясь, достигают максимума развития в разгар лучевой болезни. У выживших животных происходит восстановление естественных факторов иммунитета, полнота которого определяется степенью лучевого поражения.
Следует отметить, что в отношении действия ионизирующего излучения на факторы естественного иммунитета еще много невыясненного, в частности, слабо изучены вопросы последовательности их угнетения, значение каждого из них при различных инфекциях и у разных животных, возможности их компенсации и активации.