Гистология гипофиза: строение и развитие. Гистология центральных органов эндокринной системы Аденогипофиз гистология
Читайте также
Аденогипофиз развивается из эпителия крыши ротовой полости, имеющей эктодермальное происхождение. На 4-й неделе эмбриогенеза образуется эпителиальное выпячивание этой крыши в виде кармана Ратке. Проксимальный отдел кармана редуцируется, и ему навстречу выпячивается дно 3 желудочка, из которого образуется задняя доля. Из передней стенки кармана Ратке образуется передняя доля, из задней — промежуточная. Соединительная ткань гипофиза формируется из мезенхимы.
Функции гипофиза:
регуляция деятельности аденогипофиззависымых эндокринных желез;
накопление для нейрогормонов гипоталамуса вазопрессина и окситоцина;
регуляция пигментного и жирового обмена;
синтез гормона, регулирующего рост организма;
выработка нейропептидов (эндорфинов).
Гипофиз представляет собой паренхиматозный орган со слабым развитием стромы. Он состоит из аденогипофиза и нейрогипофиза. Аденогипофиз включает три части: переднюю, промежуточную доли и туберальную часть.
Передняя доля состоит из эпителиальных тяжей трабекул, между которыми проходят фенестрированные капилляры. Клетки аденогипофиза называются аденоцитами. В передней доле их 2 вида.
Хромофильные аденоциты располагаются по периферии трабекул и содержат в цитоплазме гранулы секрета, которые интенсивно окрашиваются красителями и делятся на: оксифильные и базофильные.
Оксифильные аденоциты делятся на две группы:
соматотропоциты вырабатывают гормон роста (соматотропин), стимулирующий деление клеток в организме и его рост;
лактотропоциты вырабатывают лактотропный гормон (пролактин, маммотропин). Этот гормон усиливает рост молочных желез и секрецию ими молока во время беременности и после родов, а также способствует образованию в яичнике желтого тела и выработке им гормона прогестерона.
Базофильные аденоциты подразделяются также на два вида:
тиротропоциты — вырабатывают тиреотропный гормон, этот гормон стимулирует выработку щитовидной железой тиреоидных гормонов;
гонадотропоциты подразделяются на два вида — фоллитропоциты вырабатывают фолликулостимулирующий гормон, в женском организме он стимулирует процессы овогенеза и синтез женских половых гормонов эстрогенов. В мужском организме фолликулостимулирующий гормон активирует сперматогенез. Лютропоциты вырабатывают лютеотропный гормон, который в женском организме стимулирует развитие желтого тела и секрецию им прогестерона.
Еще одна группа хромофильных аденоцитов — адренокортикотропоциты. Они лежат в центре передней доли и вырабатывают адренокортикотропный гормон, стимулирующий секрецию гормонов пучковой и сетчатой зонами коры надпочечников. Благодаря этому адренокортикотропный гормон участвует в адаптации организма к голоданию, травмам, другим видам стресса.
Хромофобные клетки сосредоточены в центре трабекул. Эта неоднородная группа клеток, в которой выделяют следующие разновидности:
незрелые, малодифференцированные клетки, играющие роль камбия для аденоцитов;
выделившие секрет и потому не окрашивающиеся в данный момент хромофильные клетки;
фолликулярно-звездчатые клетки — небольших размеров, имеющие небольшие отростки, при помощи которых они соединяются друг с другом и образуют сеть. Функция их не ясна.
Средняя доля состоит из прерывистых тяжей базофильных и хромофобных клеток. Имеются кистозные полости, выстланные реснитчатым эпителием и содержащие коллоид белковой природы, в котором отсутствуют гормоны. Аденоциты промежуточной доли вырабатывают два гормона:
меланоцитостимулирующий гормон, он регулирует пигментный обмен, стимулирует выработку меланина в коже, адаптирует сетчатку в видению в темноте, активирует кору надпочечников;
липотропин, который стимулирует жировой обмен.
Туберальная зона образована тонким тяжом эпителиальных клеток, окружающих эпифизарную ножку. В туберальной доле проходят гипофизарные портальные вены, соединяющие первичную капиллярную сеть медиального возвышения с вторичной капиллярной сетью аденогипофиза.
Задняя доля или нейрогипофиз имеет нейроглиальное строение. В ней гормоны не вырабатываются, а лишь накапливаются. Сюда поступают по аксонам и депонируются в тельцах Геринга вазопрессин и окситоциннейрогормоны переднего гипоталамуса. Состоит нейрогипофиз из эпендимных клеток — питуицитов и аксонов нейронов паравентрикулярных и супраоптических ядер гипоталамуса, а также кровеносных капилляров и телец Геринга — расширений аксонов нейросекреторных клеток гипоталамуса. Питуициты занимают до 30 % объема задней доли. Они имеют отростчатую форму и образуют трехмерные сети, окружая аксоны и терминали нейросекреторных клеток. Функциями питуицитов является трофическая и поддерживающая функции, а также регуляция выделения нейросекрета из терминалей аксонов в гемокапилляры.
Кровоснабжение аденогипофиза и нейрогипофиза изолированное. Аденогипофиз кровоснабжается из верхней гипофизарной артерии, которая вступает в медиальную эминенцию гипоталамуса и распадается на первичную капиллярную сеть. На капиллярах этой сети заканчиваются аксовазальными синапсами аксоны нейросекреторных нейронов медиобазального гипоталамуса, вырабатывающих рилизинг-факторы. Капилляры первичной капиллярной сети и аксоны вместе с синапсами образуют первый нейрогемальный орган гипофиза. Затем капилляры собираются в портальные вены, которые идут в переднюю долю гипофиза и там распадаются на вторичную капиллярную сеть фенестрированного или синусоидного типа. По ней рилизинг-факторы достигают аденоцитов и сюда же выделяются гормоны аденогипофиза. Эти капилляры собираются в передние гипофизарные вены, которые несут кровь с гормонами аденогипофиза к органам-мишеням. Поскольку капилляры аденогипофиза лежат между двумя венами (портальной и гипофизарной), они относятся к "чудесной" капиллярной сети. Задняя доля гипофиза кровоснабжается нижней гипофизарной артерией. Эта артерия распадается до капилляров, на которых образуются аксовазальные синапсы нейросекреторных нейронов — второй нейрогемальный орган гипофиза. Капилляры собираются в задние гипофизарные вены.
1. Основные этапы становления гемацитопоэза и иммуноцитопоэза в филогенезе.
2. Классификация органов кроветворения.
3. Общая морфофункциональная характеристика органов кроветворения. Понятие о специфическом микроокружении в органах кроветворения.
4. Красный костный мозг: развитие, строение и функции.
5. Тимус - центральный орган лимфоцитопоэза. Развитие, строение и функции. Возрастная и акцидентальная инволция тимуса.
В процессе эволюции происходит изменение топографии органов кроветворения (ОКТ), усложнение их строения и дифференциация функций.
1. У беспозвоночных: нет еще четкой органной локализации кроветворной ткани; примитивные клетки гемолимфы (амебоциты) рассеяны по тканям органов диффузно.
2. У низших позвоночных (круглоротые): появляются первые обособленные очаги кроветворения в стенке пищеварительной трубки. Основу этих очагов кроветворения составляет ретикулярная ткань, имеются синусоидные капилляры.
3. У хрящевых и костистых рыб наряду с очагами кроветворения в стенке пищеварительной трубки появляются обособленные ОКТ - селезенка и тимус; имеются очаги КТ и в гонадах, интерреналевых тельцах и даже в эпикарде.
4. У высокоорганизованных рыб впервые очаги КТ появляются в костной ткани.
5. У земневодных происходит органное разделение миелопоэза и лимфопоэза.
6. У пресмыкающихся и птиц четкое органное разделение миелоидной и лимфоидной ткани; основной ОКТ - красный костный мозг.
7. У млекопитающих - основной ОКТ - красный костный мозг, в остальных органах - лимфоцитопоэз.
Классификация ОКТ:
I. Центральные ОКТ
1. Красный костный мозг
II. Периферические ОКТ
1.Собственно лимфоидные органы (по ходу лимфатических сосудов - лимфатические узлы).
2. Гемолимфоидные органы (по ходу кровеносных сосудов - селезенка, гемолимфатические узлы).
3. Лимфоэпителиальные органы (лимфоидные скопления под эпителием слизистых оболочек пищеварительной, дыхательной, мочеполовой системы).
Общая морфофункциональная характеристика ОКТ
Несмотря на значительное разнообразие ОКТ имеют много общего - в источниках развития, в строении и функциях:
1. Источник развития - все ОКТ закладываются из мезенхимы; исключением является тимус - развивается из эпителия 3-4-ых жаберных карманов.
2. Общность в строении - основу всех ОКТ составляет соединительная ткань со специальными свойствами - ретикулярная ткань. Исключение - тимус: основу этого органа составляет сетчатый эпителий (ретикулоэпителиальная ткань).
3. Кровоснабжение ОКТ - обильно кровоснабжаются; имеют гемокапилляры синусоидного типа (диаметр 20 и более мкм; между эндотелиоцитами имеются большие щели, поры, базальная мембрана не сплошная - местами отсутствует; кровь протекает медленно).
Роль ретикулярной ткани в ОКТ
Вы помните, что РТ состоит из клеток (ретикулярнык клетки, в небольшом количестве фибрабластоподобные клетки, макрофаги, тучные и плазматические клетки, остеогенные клетки) и межклеточного вещества, представленное ретикулярными волокнами и основным аморфным веществом. Ретикулярная ткань в ОКТ выполняет функции:
1. Создает специфическое микроокружение, определяющее направление дифференцировки созревающих клеток крови.
2. Трофика созревающих клеток крови.
3. Фагоцитоз и утилизация погибших клеток крови за счет фагоцитоза ретикулярных клеток и макрофагов.
4. Опорно-механическая функция - является несущим каркасом для созревающих клеток крови.
КРАСНЫЙ КОСТНЫЙ МОЗГ - центральный ОКТ, где идет как миелопоэз, так и лимфоцитопоэз. ККМ в эмбриональном периоде закладывается из мезенхимы на 2-ом месяце, к 4-му месяцу становится центром кроветворения. ККМ - ткань полужидкой консистенции, темно-красного цвета из-за большого содержания эритроцитов. Небольшое количество ККМ для исследований можно получить путем пункции грудины или гребня подвздошной кости.
Строму ККМ составляет ретикулярная ткань, обильно пронизанная гемокапиллярами синусоидного типа. В петлях ретикулярной ткани располагаются островками или колониями созревающие клетки крови:
1. Эритроидные клетки в своих островках-колониях сгруппируются вокруг макрофагов, нагруженных железом, полученных от погибших в селезенке старых эритроцитов. Макрофаги в ККМ передают эритроидным клеткам железо, необходимое для синтеза ими гемоглобина.
2. Отдельными островками-колониями вокруг синусоидных гемокапилляров располагаются лимфоциты, гранулоциты, моноциты, мегакариоциты. Островки разных ростков перемежаются друг с другом и создают мозаичную картину.
Созревшие клетки крови проникают через стенки в синусоидные гамокапилляры и уносятся кровотоком. Прохождению клеток через стенки сосудов способствует повышенная проницаемость синусоидных гемокапилляров (щели, отсутствие местами базальной мембраны), высокое гидростатическое давление в ретикулярной ткани органа. Высокое гидростатическое давление обусловлено 2-мя обстоятельствами:
1. Клетки крови размножаются в ограниченной костной тканью замкнутом пространстве, обьем которого не может меняться и это приводит к повышению давления.
2. Суммарный диаметр приносящих сосудов больше диаметра выносящих сосудов, что тоже приводит к повышения давления.
Возрастные особенности ККМ: У детей ККМ заполняет и эпифизы и диафизы трубчатых костей, губчатое вещество плоских костей. У взрослых в диафизе ККМ замещается желтым костным мозгом (жировой тканью), а в старости студенистым костным мозгом.
Регенерация: физиологическая - за счет клеток 4-5 класса; репаративная - 1-3 классов.
ТИМУС - центральный орган лимфоцитопоэза и иммуногенеза. Тимус закладывается в начале 2-го месяца эмбрионального развития из эпителия 3-4-х жаберных карманов как экзокринная железа. В дальнейшем тяж соединяющий железу с эпителием жаберных карманов подвергается обратному развитию. В конце 2-го месяца орган заселяется лимфоцитами.
Строение тимуса - снаружи орган покрыт сдт капсулой, от которой внутрь отходят перегородки из рыхлой сдт и делят орган на дольки. Основу паренхимы тимуса составляет сетчатый эпителий: эпителиальные клетки отросчатые, соединяются друг с другом отростками и образуют петлистую сеть, в петлях которой располагаются лимфоциты (тимоциты). В центральной части дольки стареющие эпителиальные клетки образуют слоистые тимусные тельца или тельца Гассаля - концентрически наслоенные эпителиальные клетки с вакуолями, гранулами кератина и фибриллярными волокнами в цитоплазме. Количество и размеры телец Гассаля с возрастом увеличивается. Функция сетчатого эпителия:
1. Создает специфическое микроокружение для созревающих лимфоцитов.
2. Синтез гормона тимозина, необходимого в эмбриональном периоде для нормальной закладки и развития периферических лимфоидных органов, а в постнатальном периоде для регуляции функцией периферических лимфоидных органов; синтез инсулиноподобного фактора, фактора роста клеток, кальцитониноподобный фактор.
3. Трофическая - питание созревающих лимфоцитов.
4. Опорно-механическая функция - несущий каркас для тимоцитов.
В петлях сетчатого эпителия располагаются лимфоциты (тимоциты), особенно их много по периферии дольки, поэтому эта часть дольки темнее и называется корковой частью. Центр дольки содержит меньше лимфоцитов, поэтому эта часть светлее и называется мозговой частью дольки. В корковом веществе тимуса происходит "обучение" Т-лимфоцитов, т.е. они приобретают способность распознавать "свое" или "чужое". В чем суть этого обучения? В тимусе образуются лимфоциты строго специфичные (имеющие строго комплементарные рецепторы) для всех возможно мыслимых А-генов, даже против своих клеток и тканей, но в процессе "обучения" все лимфоциты имеющие рецепторы к своим тканям уничтожаются, оставляются только те лимфоциты, которые направлены против чужеродных Антигенов. Вот поэтому в корковом веществе наряду с усиленным размножением видим и массовую гибель лимфоцитов. Таким образом в тимусе из предшественников Т-лимфоцитов образуются субпопуляции Т-лимфоцитов, которые в последующем попадают в периферические лимфоидные органы, дозревают и функционируют.
После рождения масса органа в течении первых 3-х лет быстро увеличивается, медленный рост продолжается до возраста полового созревания, после 20 лет паренхима тимуса начинает замещаться жировой тканью, но минимальное количество лимфоидной ткани сохраняется до глубокой старости.
Акцидентальная инволюция тимуса (АИТ): Причиной акцидентальной инволюции тимуса могут быть чрезмерно сильные раздражители (травма, инфекции, интоксикации, сильные стрессы и т.д.). Морфологически АИТ сопровождается массовой миграцией лимфоцитов из тимуса в кровоток, массовой гибелью лимфоцитов в тимусе и фагоцитозом погибших клеток макрофагами (иногда фагоцитоз и нормальных, не погибших лимфоцитов), разрастанием эпителиальной основы тимуса и усилением синтеза тимозина, стиранием границы между корковой и мозговой частью долек. Биологичесое значение АИТ:
1. Гибнущие лимфоциты являются донорами ДНК, которая транспортируется макрофагами в очаг поражения и используется там пролиферирующими клетками органа.
2. Массовая гибель лимфоцитов в тимусе является проявлением селекции и элиминации Т-лимфоцитов, имеющих рецепторы против собственных тканей в очаге поражения и направлена на предотвращение возможной аутоагрессии.
3. Разрастание эпителиальнотканной основы тимуса, усиление синтеза тимозина и других гормоноподобных веществ направлены на повышение функциональной активности периферических лимфоидных органов, усилению метаболических и регенераторных процессов в пораженном органе.
На препарате при малом увеличении микроскопа отчетливо видны все три доли гипофиза: передняя, промежуточная и задняя. Передняя и промежуточная доли разделены щелью, которая представляет собой остаток полости эмбрионального кармана гипофиза (карман Ратке). В ряде случаев полость наблюдается в пределах задней доли, однако это полость воронки гипофиза, связывающей его с основанием головного мозга. Передняя доля состоит из тяжей клеток, расположенных вокруг синовиальных капилляров. В пределах тяжей различаются более мелкие главные (хромофобные) клетки и более крупные хромофильные.
Промежуточная доля представляет собой скопление довольно тесно прилежащих друг к другу однородных клеток интермедиоцитов, лежащих в несколько рядов.
Задняя доля бедна клеточными элементами и состоит в основном из волокон, между которыми располагаются синусоидные капилляры и отростчатой формы клетки – питуициты.
Особенность гипофиза кошки – не зарастает карман Ратке.
Препарат № 82: Гипофиз человека (задняя доля).
Окраска: по Маллори.
чел. кош. чел.
На некоторых препаратах только передняя доля гипофиза, задней доли – нет.
 ãèïîôèçå ÷åëîâåêà íå îòìå÷àåòñÿ ñòîëü ÷åòêîãî äåëåíèÿ íà äîëè. Áîëüøóþ ÷àñòü ïðåïàðàòà çàíèìàåò ïåðåäíÿÿ äîëÿ. Ñîåäèíèòåëüíîòêàííûå âîëîêíà îêðàøåíû â èíòåíñèâíî ñèíèé öâåò è çàïîëíÿþò ïðîìåæóòêè ìåæäó òÿæàìè ýïèòåëèàëüíûõ êëåòîê.  ýòèõ ñîåäèíèòåëüíîòêàííûõ ïðîñëîéêàõ ðàñïîëàãàþòñÿ ìíîãî÷èñëåííûå ñèíóñîèäíûå êàïèëëÿðû, ñîäåðæàùèå ýëåìåíòû êðîâè. Ïðè áîëüøîì óâåëè÷åíèè âèäíî, ÷òî áîëüøóþ ÷àñòü êëåòîê ïàðåíõèìû ñîñòàâëÿþò õðîìîôîáíûå àäåíîöèòû – ìåëêèå êëåòêè, êîòîðûå áëåäíî îêðàøèâàþòñÿ êèñëûìè êðàñèòåëÿìè. Âòîðîé òèï êëåòîê – àöèäîôèëüíûå àäåíîöèòû – îòëè÷àþòñÿ îò õðîìîôîáíûõ áîëåå êðóïíûìè ðàçìåðàìè è áîëåå îêñèôèëüíîé öèòîïëàçìîé. È íàêîíåö, ñàìàÿ ìàëî÷èñëåííàÿ ãðóïïà – áàçîôèëüíûå àäåíîöèòû – êðóïíûå êëåòêè ñ áàçîôèëüíîé îêðàñêîé öèòîïëàçìû.
Препарат № 83: Щитовидная железа собаки.
Окраска: гематоксилин-эозин.
При малом увеличении микроскопа можно увидеть, что железа покрыта снаружи соединительнотканной капсулой и разделена прослойкой соединительной ткни на дольки различной величины. В междольковой соединительной ткани можно встретить сосуды: артерии и вены. Дольки состоят из округлой формы фолликулов, тесно прилежащих друг к другу. Каждый фолликул окружен тонкой прослойкой соединительной ткани, в которой располагаются многочисленные капилляры.
При большом увеличении видно, что стенка фолликула образована одним слоем клеток – фолликулярных тиреоцитов (форма клеток варьирует в зависимости от функционального состояния железы). Просвет фолликула заполнен коллоидом – гомогенной оксифильной массой. Второй вид клеточных элементов железы – парафолликулярные тиреоциты – может располагаться как в пределах стенки фолликула, так и рядом с ним парафолликулярно, но они не соприкасаются с коллоидом, отделенные от него узкими участком цитоплазмы фолликулярного тиреоцита. При окраске гематоксилин-эозином эти клетки с трудом дифференцируются от фолликулярных тиреоцитов. Между фолликулами встречаются не имеющие полости скопления эпителиальных клеток – интерфолликулярные островки, - состоящие из крупных клеток со светлой цитоплазмой.
ОТЛИЧИЯ ЩИТОВИДНОЙ ЖЕЛЕЗЫ НОВОРОЖДЕННОГО ОТ ВЗРОСЛОГО:
Ïðè áîëüøîì óâåëè÷åíèè ìèêðîñêîïà – ìåíüøèé ðàçìåð ôîëëèêóëîâ è áîëüøåå, ÷åì ó âçðîñëîãî, êîëè÷åñòâî ìåæôîëëèêóëÿðíûõ îñòðîâêîâ. Êîëëîèä îêðàøèâàåòñÿ ñëàáåå âñëåäñòâèå ìåíüøåé ïëîòíîñòè. Çíà÷èòåëüíî ðàñøèðåííûå êàïèëëÿðû, ðàñïîëàãàþòñÿ â ïðîñëîéêàõ ñîåäèíèòåëüíîé òêàíè ìåæäó ôîëëèêóëàìè.
Препарат № 84: Паращитовидная железа быка.
Окраска: гематоксилин-эозин.
Парафолликулярные тиреоциты отличаются от фолликулярных тем, что их цитоплазма интенсивно окрашивается солями серебра. Более крупные, чем фолликулярные, они располагаются одиночно или группами по 2-3 клетки в составе стенки фолликула, отделенные от его просвета участком цитоплазмы фолликулярного тиреоцита или в составе интерфолликулярных островков.
Каждая железа представляет собой небольшое эпительальное тельце, включенное в ткань щитовидной железы, поэтому в препарате могут встречаться участки щитовидной железы.
Железа состоит из различной формы и величины извитых тяжей эпителиальных клеток – паратиреоцитов, разделенных прослойками рыхлой соединительной ткани, иногда содержащей большое количество жировых клеток. В прослойках соединительной ткани располагаются многочисленные капилляры.
Препарат № 85: Надпочечник собаки (или взрослого).
Окраска: железный гематоксилин.
При малом увеличении микроскопа – орган сверху покрыт капсулой, содержащей жировые клетки и крупные сосуды. Под капсулой находится корковое вещество, разделенное на три зоны, отличающиеся характером расположения эпителиальных тяжей – в самой верхней (клубочковой) зоне они образуют округлой формы скопления, затем идет пучковая зона, где клетки лежат параллельными тяжами, и наконец, в сетчатой зоне тяжи переплетаются друг с другом, наподобие сети. Мозговое вещество нечетко отграничено от коркового тонкой прослойкой соединительной ткани и представлено более крупными клетками, чем клетки сетчатой зоны. Для мозгового вещества характерно наличие крупных расширенных синусоидных капилляров. Кроме того, капилляры пронизывают как мозговое, так и корковое вещество, располагаясь в соединительнотканных прослойках.
При большом увеличении можно рассмотреть элементы, входящие в состав органа. Клетки мозгового вещества содержат в цитоплазме зернистость, представляющую собой скопление секреторного продукта.
Препарат № 87: Надпочечник новорожденного.
Окраска: гематоксилин-эозин.
Ряд особенностей:
Прежде всего большее, чем у взрослого, размеры органа, которые объясняются наличием в органе широкой зоны, называемой фетальной или зародышевой корой. Она расположена между довольно узкой полоской окончательной или постоянной коры и мозговым веществом. Зародышевая кора состоит из крупных клеток, расположенных в виде тяжей, часть из которых находится в состоянии деструкции, так как непосредственно перед рождением и в первые недели после него происходит интенсивная гибель клеток фетальной коры. Сосуды, расположенные в этой зоне, расширены и заполнены кровью, вследствие чего эта зона может быть хорошо дифференцирована от остальных частей органа.
В постоянном корковом веществе надпочечника не представляется возможным отдифференцировать образующие его зоны.
Мозговое вещество занимает значительно меньший объем, чем у взрослого, располагается в центре железы и может встречаться в виде «мозговых шаров» по всему надпочечнику. Мозговые шары, представляющие собой скопление малодифференцированных симпатогоний, мигрируют к центру железы. В процессе миграции идет дифференцировка симпатогоний на симпатобласты и хромоффинобласты.
расширенные большие сосуды
могут быть видны последние «мозговые шары»
(ближе к центру) и мозговое вещество.
Препарат № 88: Желтое тело беременности из яичника свиньи.
Окраска: гематоксилин-эозин.
Желтое тело – это временный инкреторный орган, образовавшийся на месте фолликула после овуляции. Препарат представляет собой срез желтого тела в стадии расцвета. Структуру желтого тела следует рассмотреть под большим увеличением. Желтое – это накопившийся прогестерон.
Основу желтого тела составляют крупные светлые лютеиновые клетки (лютеоциты), представляющие собой гиперторофированные клетки бывшего зернистого слоя фолликула, содержащие желтый пигмент лютеин, относящийся к группе липохромов. Лютеиноциты разделены тончайшими прослойками соединительной ткани, сопровождающими капилляры.
Препарат № 89: Островки Лангерганса (поджелудочная железа плода).
Окраска: гематоксилин-эозин.
При малом увеличении микроскопа видно, что поджелудочная железа разделена соединительнотканными прослойками на дольки. Основная масса долек представлена концевыми секреторными отделами (экзокринная часть железы - сложная альвеолярно-трубчатая, белковая) – ацинусами, между которыми имеются светлые вкрапления – островки Лангерганса (эндокринная часть железы). В междольковой соединительной ткани видны междольковые выводные протоки, выстланные однослойным призматическим эпителием, сосуды (артерии, вены), нервные стволы, интрамуральные нервные узлы. При большом увеличении микроскопа отчетливо выявляется структурная организация концевых отделов экзокринной части железы. Они имеют небольшой просвет, образованы эпителиальными клетками конической формы, цитоплазма которых подразделена на гомогенную (темную базальную) зону и зимогенную (светлую апикальную). Ядра секреторных клеток располагаются центрально. Система выводных протоков начинается со вставочного отдела, который достигает значительной длины и образован плоскими эпителиальными клетками, лежащими на базальной мембране. Вставочный проток постепенно переходит во внутридольковый небольшой протяженности, выстланный кубическим эпителием, который переходит затем в междольковый проток. Панкреатические островки образованы связанными между собой тяжами или компактными группами светлых клеток полигональной формы, между которыми залегают синусоидные капилляры. При помощи специальных методов окрашивания в панкреатических островках можно выделить несколько типов клеток.
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА ЭМБРИОНА 8 НЕДЕЛЬ.
Ïðè ìàëîì óâåëè÷åíèè ìèêðîñêîïà âèäíû íåïîñðåäñòâåííî ïîä æåëóäêîì ôîðìèðóþùèåñÿ òðóá÷àòûå îáðàçîâàíèÿ (âûâîäíûå ïðîòîêè), ðàçäåëåííûå ïðîñëîéêàìè çàðîäûøåâîé ñîåäèíèòåëüíîé òêàíè, çàíèìàþùåé çíà÷èòåëüíîå ìåñòî â çàêëàäêå æåëåçû. Äèôôåðåíöèðîâêà ñåêðåòîðíûõ êëåòîê íà÷èíàåòñÿ íåñêîëüêî ïîçäíåå (ñ 10-11 íåäåëè), à ïåðâûå àöèíóñû ïîÿâëÿþòñÿ â 12-14 íåäåëü.
ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА НОВОРОЖДЕННОГО.
В первые дни после рождения железа характеризуется незрелостью основных своих компонентов: дольки некомпактны, центральная часть дольки занята стромой, которая очень обильна в железе. Четкого деления на дольки не отмечается. Концевые отделы состоят из клеток небольшого размера, цитоплазма которых, однако, уже дифференцирована на гомогенную и зимогенную зону. Островковая часть железы развита хорошо и практически
идентична таковому аппарату взрослого человека.
Дата добавления: 2015-05-19 | Просмотры: 1124 | Нарушение авторских прав
1 | | | | | | | | | | | |
Материал взят с сайта www.hystology.ru
Гипофиз - компонент единой гипоталамофизарной системы организма. Вырабатывает гормоны, регулирующие функцию многих желез внутренней секреции и осуществляет их связь с центральной нервной системой. Расположен он в гипофизарной ямке турецкого седла клиновидной кости черепа; имеет бобовидную форму и очень небольшую массу. Так, у крупного рогатого скота она около 4 г, а у свиней меньше - 0,4 г.
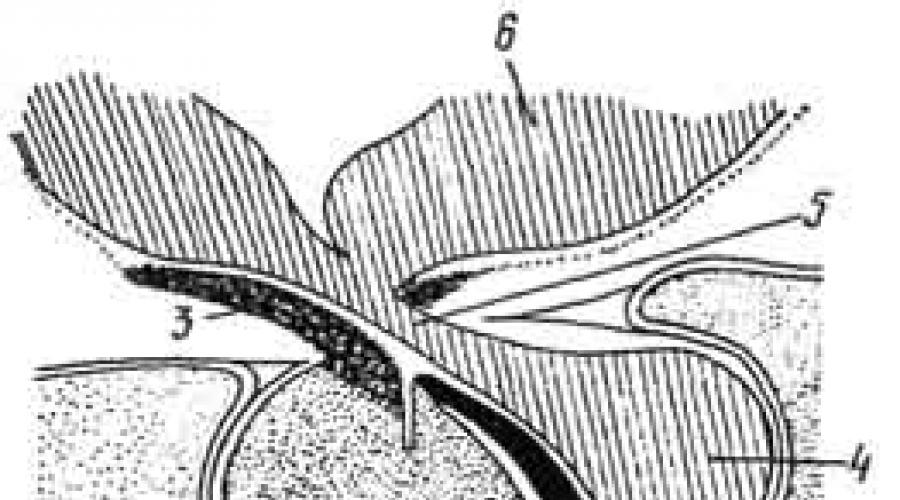
Развивается гипофиз из двух эмбриональных зачатков, растущих навстречу друг другу. Первый зачаток - гипофизарный карман - образуется из крыши первичной ротовой полости и направлен в сторону головного мозга. Это эпителиальный зачаток, из которого в дальнейшем развивается аденогипофиз.
Второй зачаток - выпячивание дна мозгового желудочка, поэтому он является мозговым карманом и из него образуется нейрогипофиз (рис. 217).
Эмбриогенез обусловил строение органа - гипофиз состоит из двух долей: аденогипофиза и нейрогипофиза (рис. 218, 219).
В состав аденогипофиза входят передняя, промежуточная и туберальная части. Передняя часть построена из эпителиальных клеток - аденоцитов, формирующих тяжи (трабекулы) и разграниченных синусоидными капиллярами вторичной сосудистой сети (цв. табл. VII - А - а). Первичная сосудистая сеть расположена в медиальной эминенции.
Рис. 217. Развитие гипофиза:
А - ранняя и Б - более поздняя стадии; а - стенки нервной трубки; б - стенки мозгового пузыря; в - эпителий ротовой бухты; г - хорда; д - кишечная трубка; е - мезенхима; ж - гипофизарный карман; его з - передняя и и - задняя стенки; к - зачаток задней части.

Рис. 218. Строение гипофиза:
1 - передняя, 2 - промежуточная, 3 - туберальная и 4 - задняя части; 5 - воронка; 6 - гипоталамус.

Рис. 219. Схема медиального разреза гипофиза домашних животных:
а - лошади; б - крупного рогатого скота; в - свиньи; г - собаки; д - кошки (по Траутману и Фибигеру).
Соединительнотканная строма аденогипофиза развита слабо.
Аденоциты по-разному воспринимают красители: клетки, хорошо окрашивающиеся, называют хромофильными, а плохо красящиеся - хромофобными (б). Хромофильные аденоциты могут воспринимать или кислые красители или основные, поэтому первые именуют ацидофильными (б), вторые - базофильными (г).
Ацидофильные клетки составляют 30 - 35% всех клеток передней части гипофиза. Они имеют округлую или овальную форму, крупнее хромофобных и мельче базофильных аденоцитов. В цитоплазме ацидофила содержатся гранулы, окрашивающиеся эозином; ядро расположено в центре клетки. К нему прилегают комплекс Гольджи, незначительное количество крупных митохондрий, хорошо развита зернистая эндоплазматическая сеть, что свидетельствует об интенсивном синтезе белка.
В связи с разной гормонообразовательной функцией и строением, цитоплазматической зернистостью различают три разновидности ацидофильных аденоцитов: соматотропоциты, лактотропоциты, кортикотропоциты. Соматотропоциты продуцируют соматотропный гормон, стимулирующий рост тканей и всего организма в целом. Лактотропоциты образуют пролактин (лактотропный гормон), регулирующий процесс лактации и функциональное состояние желтого тела яичника. Кортикотропоциты вырабатывают кортикотропин, повышающий гормонообразовательную функцию коры надпочечников.
Секреторные гранулы соматотропоцитов шаровидной формы, в диаметре от 200 до 400 нм (рис. 220). Лактотропоциты имеют более крупные секреторные гранулы овальной формы с длиной 500 - 600 нм, шириной 100 - 120 нм. Секреторные гранулы кортикотропоцитов снаружи покрыты мембраной пузырчатой формы с плотной сердцевиной.
Базофильные аденоциты составляют 4 - 10% всех клеток передней части гипофиза. Это самые крупные клетки аденогипофиза. Их секреторные гранулы имеют гликопротеидный характер, поэтому окрашиваются основными красителями. Различают две разновидности этих клеток: гонадотропные и тиротропные. Гонадотропные клетки продуцируют фолликулостимулирующий гормон, регулирующий развитие женских и мужских половых клеток, секрецию половых органов самки и лютеинизирующего гормона, стимулирующего рост и развитие желтого тела в яичниках и

Рис. 220. Соматотропоцит передней доли аденогипофиза (электронная микрофотография):
1 - гранулярная эндоплазматическая сеть; 2 - комплекс Гольджи; 3 - Формирующиеся гранулы секрета; 4 - ядро; 5 - зрелые гранулы секрета; 6 - ? ;митохондрия (по Стрижкову).

Рис. 221. Гонадотропоцит передней доли аденогипофиза:
1 - ядро; 2 - комплекс Гольджи; 3 - секреторные гранулы; 4 - накопительные гранулы; 5 - митохондрии; 6 - цистерны гранулярной эндоплазматической сети.
интерстициальных клеток в семенниках (рис. 221). В центральной зоне гонадотропного базофила находится макула. Это расширенная полость комплекса Гольджи, оттесняющая ядро, многочисленные мелкие митохондрии, мембраны эндоплазматической сети на: периферию клетки. Базофильные гонадотропоциты содержат зернистость, равную около 200 - 300 нм в диаметре.
При недостаточности половых гормонов в организме диаметр зернистости увеличивается. После кастрации животных базофильные гонадотропоциты превращаются в клетки кастрации: крупная вакуоль занимает всю центральную часть клетки. Последняя приобретает кольцевидную форму.
Тиреотропные базофилы (рис. 222) - угловатые клетки с мелкой (80 - 150 нм) заполняющей всю цитоплазму зернистостью. Если

Рис. 222. Тиреотропоцит передней доли аденогипофиза (электронная микрофотография):
1 - ядро; 2 - секреторные гранулы; 3 - соматотропоцит (по Долан и Сёлоши).
организм испытывает недостаток гормонов щитовидной железы, то развиваются клетки тиреодэктомии. Они увеличены в размере, с расширенными цистернами эндоплазматической сети, поэтому цитоплазма имеет ячеистый вид, более крупные гранулы секрета,
Хромофобные клетки составляют 60 - 70% всех клеток передней части гипофиза. Это сборная группа, так как в ее состав входят разные по значению клетки: камбиальные, клетки на разной стадии дифференциации; не накопившие еще специфическую зернистость; клетки, выделившие секрет. Из камбиальных клеток в дальнейшем развиваются ацидофильные и базофильные аденоциты.
Промежуточная часть аденогипофиза представлена несколькими рядами слабобазофильных клеток. Продуцируемый
аденоцитами секрет накапливается в промежутках между клетками, что способствует образованию фолликулоподобных структур. Клетки промежуточной части аденогипофиза полигональной формы, содержат мелкие гликопротеидные гранулы размером 200 - 300 ни. В промежуточной зоне синтезируются меланотропин, регулирующий пигментный обмен, и липотропин - стимулятор жирового обмена.
Туберальная часть аденогипофиза по своей структуре сходна с промежуточной частью. Она прилегает к гипофизарной ножке и медиальной эминенции. Клетки этой зоны характеризуются слабой базофилией и трабекулярным расположением. Функция туберальной части окончательно не выяснена.
Выше говорилось о том, что гормонообразовательная функция аденогипофиза регулируется гипоталамусом, с которым он образует единую гипоталамоаденогипофизарную систему. Морфофункционально эта связь проявляется в следующем: верхняя гипофизарная артерия в медиальной эминенции образует первичную капиллярную сеть. Аксоны мелких нейросекреторных клеток ядер медиобазального гипоталамуса на сосудах первичной капиллярной сети образуют аксоваскулярные синапсы. Нейрогормоны, продуцируемые этими нейросекреторными клетками, по их аксонам перемещаются в медиальную эминенцию. Здесь они накапливаются, а затем через аксоваскулярные синапсы поступают в капилляры первичной сосудистой сети. Последние собираются в портальные вены, которые направляются вдоль гипофизарной ножки в аденогипофиз. Здесь вновь они распадаются и образуют вторичную капиллярную сеть. Синусоидные капилляры этой сети оплетают трабекулы секретирующих аденоцитов.
Кровь, оттекающая по венам от вторичной сосудистой сети, содержит аденогипофизарные гормоны, которые через общий кровоток, то есть гуморальным способом, регулируют функции эндокринных желез периферического звена.
Нейрогипофиз (задняя доля) развивается из мозгового кармана, поэтому он построен из нейроглии. Его клетки - питуициты веретенообразной или отростчатой формы. Отростки питуицитов контактируют с кровеносными сосудами. В заднюю долю входят крупные пучки нервных волокон, образованные аксонами нейросекреторных клеток паравентрикулярных и супраоптических ядер передней зоны гипоталамуса. Нейросекрет, образовавшийся этими клетками, перемещается вдоль аксонов в нейрогипофиз в виде секреторных капель. Здесь они оседают в виде накопительных телец, или терминалей, которые контактируют с капиллярами.
Следовательно, гормоны нейрогипофиза - окситоцин и вазопрессин синтезируются не структурами нейрогипофиза, а в паравентрикулярных и супраоптических ядрах. Затем, как говорилось выше, по нервным волокнам гормоны поступают в нейрогипофиз, где они накапливаются и откуда поступают в кровяное русло. Поэтому нейрогипофиз и гипоталамус тесно связаны и формируют единую гипоталамонейрогипофизарную систему.
Окситоцин стимулирует функцию гладких мышц матки, способствует выделению секрета маточных желез; при родах вызывает сильное сокращение мышечной оболочки стенки матки; регулирует сокращение мышечных элементов молочной железы.
Вазопрессин суживает просвет кровеносных сосудов и повышает кровяное давление; регулирует водный обмен, так как влияет на обратное всасывание (реабсорбцию) воды в канальцах почки.