Экологическая система. Естественная экосистема
Читайте также
| В агроэкосистеме преобладают растения одного вида (например, на пшеничном поле – пшеница). Растения других видов, животные и грибы присутствуют, но их меньше, чем в природной экосистеме. Низкое видовое разнообразие приводит к тому, что пищевые цепи в агроэкосистеме короткие, неразветвленные. Из-за этого круговорот веществ в агроэкосистеме неустойчивый – следовательно, и сама агроэкосистема неустойчива (ее саморегуляция ослаблена). Чтобы не разрушиться, агроэкосистема должна получать энергию не только от Солнца, но и из дополнительных источников (полив, удобрения, прополка). В естественной экосистеме растения своими корнями забирают из почвы минеральные соли, затем растения поедаются консументами, разрушаются редуцентами, и соли возвращаются назад в почву - это замкнутый круговорот веществ. На поле пшеницы урожай собирается и вывозится, и минеральные соли в почву не возвращаются - это незамкнутый круговорот веществ. |
1. Установите соответствие между характеристикой экосистем и их типом: 1-природная экосистема, 2-агроэкосистема
А) преобладают растения одного вида
Б) обитает большое разнообразие видов
В) осуществляется саморегуляция численности популяций
Г) круговорот веществ незамкнутый
Д) большую роль играет антропогенный фактор
Е) пищевые цепи длинные
Ответ
А2 Б1 В1 Г2 Д2 Е1
2. В агроэкосистеме картофельного поля, в отличие от экосистемы луга
А) отсутствуют консументы
Б) высокая численность продуцентов одного вида
В) незамкнутый круговорот веществ
Г) преобладают растительноядные насекомые
Д) отсутствуют редуценты
Е) нарушена саморегуляция
Ответ
3. В чём сходство природной и искусственной экосистем?
А) небольшое число видов
Б) наличие цепей питания
В) замкнутый круговорот веществ
Г) использование солнечной энергии
Д) использование дополнительных источников энергии
Е) наличие продуцентов, консументов, редуцентов
Ответ
4. В природной экосистеме, в отличие от искусственной
A) длинные цепи питания
Б) короткие цепи питания
B) небольшое число видов
Ответ
5. В естественной экосистеме
А) разнообразный видовой состав
Б) незначительная саморегуляция
В) численность популяций не изменяется
Г) замкнутый круговорот веществ
Д) разветвленные пищевые цепи
Е) среди консументов преобладают хищники
Ответ
6. В природной экосистеме, в отличие от искусственной
А) длинные цепи питания
Б) продуценты изымаются из круговорота
В) небольшое число видов
Г) осуществляется саморегуляция
Д) замкнутый круговорот веществ
Е) используются дополнительные источники энергии наряду с солнечной
Ответ
7. В экосистеме широколиственного леса – дубраве –
А) короткие пищевые цепи
Б) устойчивость обеспечивается разнообразием растений и животных
В) высокая биологическая продуктивность
Г) видовой состав растений и животных ограничен
Д) почва богата гумусом
Е) в почве отсутствуют редуценты
Экосистема — это функциональное единство живых организмов и среды их обитания. Основные характерные особенности экосистемы — ее безразмерность и безранговость. Замещение одних биоценозов другими в течение длительного периода времени называется сукцессией. Сукцессия, протекающая на вновь образовавшемся субстрате, называется первичной. Сукцессия на территории, уже занятой растительностью, называется вторичной.
Единицей классификации экосистем является биом — природная зона или область с определенными климатическими условиями и соответствующим набором доминирующих видов растений и животных.
Особая экосистема — биогеоценоз — участок земной поверхности с однородными природными явлениями. Составными частями биогеоценоза являются климатоп, эдафотоп, гидротоп (биотоп), а также фитоценоз, зооценоз и микробоценоз (биоценоз).
С целью получения продуктов питания человек искусственно создает агроэкосистемы. Они отличаются от естественных малой устойчивостью и стабильностью, однако более высокой продуктивностью.
Экосистемы — основные структурные единицы биосферы
Экологическая система, или экосистема, — основная функциональная единица в экологии, так как в нее входят организмы и
неживая среда — компоненты, взаимно влияющие на свойства друг друга, и необходимые условия для поддержания жизни в той ее форме, которая существует на Земле. Термин экосистема впервые был предложен в 1935 г. английским экологом А. Тенсли.
Таким образом, под экосистемой понимается совокупность живых организмов (сообществ) и среды их обитания, образующих благодаря круговороту веществ, устойчивую систему жизни.
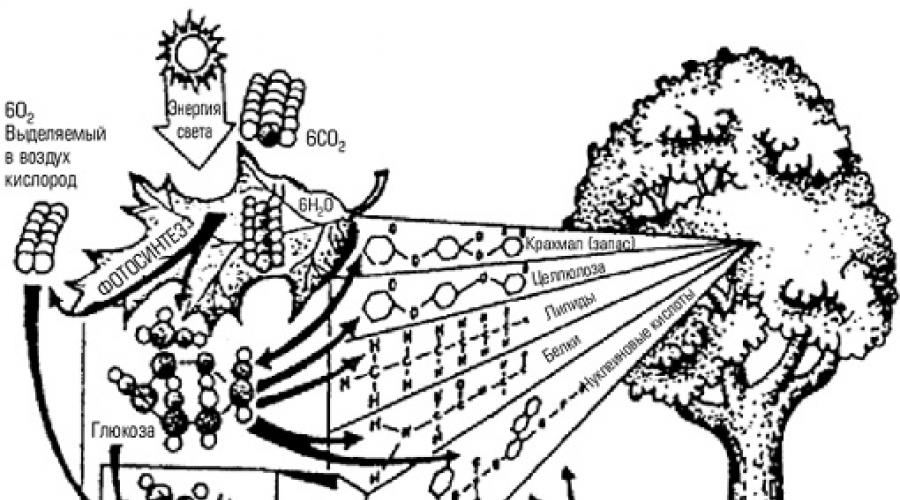
Сообщества организмов связаны с неорганической средой теснейшими материально- энергетическими связями. Растения могут существовать только за счет постоянного поступления в них углекислого газа, воды, кислорода, минеральных солей. Гетеротрофы живут за счет автотрофов, но нуждаются в поступлении таких неорганических соединений, как кислород и вода.
В любом конкретном месте обитания запасов неорганических соединений, необходимых для поддержания жизнедеятельности населяющих его организмов, хватило бы ненадолго, если бы эти запасы не возобновлялись. Возврат биогенных элементов в среду происходит как в течение жизни организмов (в результате дыхания, экскреции, дефекации), так и после их смерти, в результате разложения трупов и растительных остатков.
Следовательно, сообщество образует с неорганической средой определенную систему, в которой поток атомов, вызываемый жизнедеятельностью организмов, имеет тенденцию замыкаться в круговорот.
Рис. 8.1. Структура биогеоценоза и схема взаимодействия между компонентами
В отечественной литературе широко применяется термин «биогеоценоз», предложенный в 1940 г.B . Н Сукачевым. По его определению, биогеоценоз — «совокупность на известном протяжении земной поверхности однородных природных явлений (атмосферы, горной породы, почвы и гидрологических условий), имеющая особую специфику взаимодействий этих слагающих ее компонентов и определенный тип обмена веществом и энергией их между собой и другими явлениями природы и представляющая собой внутренне противоречивое диалектическое единство, находящееся в постоянном движении, развитии».
В биогеоценозе В.Н. Сукачев выделял два блока: экотоп — совокупность условий абиотической среды и биоценоз — совокупность всех живых организмов (рис. 8.1). Экотоп часто рассматривают как абиотическую среду, не преобразованную растениями (первичный комплекс факторов физико-географической среды), а биотоп — как совокупность элементов абиотической среды, видоизмененных средообразующей деятельностью живых организмов.
Существует мнение, что термин «биогеоценоз» в значительно большей степени отражает структурные характеристики изучаемой макросистемы, тогда как в понятие «экосистема» вкладывается, прежде всего, ее функциональная сущность. Фактически же между этими терминами различий нет.
Следует указать, что совокупность специфического физико-хи- мического окружения (биотопа) с сообществом живых организмов (биоценозом) и образует экосистему:
Экосистема = Биотоп + Биоценоз.
Равновесное (устойчивое) состояние экосистемы обеспечивается на основе круговоротов веществ (см. п. 1.5). В этих круговоротах непосредственно участвуют все составные части экосистем.
Для поддержания круговорота веществ в экосистеме необходимо наличие запаса неорганических веществ в усвояемой форме и трех функционально различных экологических групп организмов: продуцентов, консументов и редуцентов.
Продуцентами выступают автотрофные организмы, способные строить свои тела за счет неорганических соединений (рис. 8.2).

Рис. 8.2. Продуценты
Консументы - гетеротрофные организмы, потребляющие органическое вещество продуцентов или других консументов и трансформирующие его в новые формы.
Редуценты живут за счет мертвого органического вещества, переводя его вновь в неорганические соединения. Классификация эта относительная, так как и консументы, и сами продуценты выступают частично в роли редуцентов в течение жизни, выделяя в окружающую среду минеральные продукты обмена веществ.
В принципе круговорот атомов может поддерживаться в системе и без промежуточного звена — консументов, за счет деятельности двух других групп. Однако такие экосистемы встречаются скорее как исключения, например на тех участках, где функционируют сообщества, сформированные только из микроорганизмов. Роль консументов выполняют в природе в основном животные, их деятельность по поддержанию и ускорению циклической миграции атомов в экосистемах сложна и многообразна.
Масштабы экосистемы в природе весьма различны. Неодинакова также степень замкнутости поддерживаемых в них круговоротов вещества, т.е. многократность вовлечения одних и тех же элементов в циклы. В качестве отдельных экосистем можно рассматривать, например, и подушку лишайников на стволе дерева, и разрушающийся пень с его населением, и небольшой временный водоем, луг, лес, степь, пустыню, весь океан и, наконец, всю поверхность Земли, занятую жизнью.
В некоторых типах экосистем вынос вещества за их пределы настолько велик, что их стабильность поддерживается в основном за счет притока такого же количества вещества извне, тогда как внутренний круговорот малоэффективен. Таковы проточные водоемы, реки, ручьи, участки на крутых склонах гор. Другие экосистемы имеют значительно более полный круговорот веществ и относительно автономны (леса, луга, озера и т.п.).
Экосистема — практически замкнутая система. В этом состоит принципиальное отличие экосистем от сообществ и популяций, являющиеся открытыми системами, обменивающимися со средой обитания энергией, веществом и информацией.
Однако ни одна экосистема Земли не имеет полностью замкнутого круговорота, поскольку минимальный обмен массой со средой обитания все-таки происходит.
Экосистема является совокупностью взаимосвязанных энергопотребителей, совершающих работу по поддержанию ее неравновесного состояния относительно среды обитания за счет использования потока солнечной энергии.
В соответствии с иерархией сообществ жизнь на Земле проявляется и в иерархичности соответствующих экосистем. Экосистемная организация жизни является одним из необходимых условий ее существования. Как уже отмечалось, запасы биогенных элементов, необходимых для жизни организмов на Земле в целом и на каждом конкретном участке на ее поверхности, небезграничны. Лишь система круговоротов могла придать этим запасам свойство бесконечности, необходимое для продолжения жизни.
Поддерживать и осуществлять круговорот могут только функционально различные группы организмов. Функционально-экологическое разнообразие живых существ и организация потока извлекаемых из окружающей среды веществ в циклы — древнейшее свойство жизни.
С этой точки зрения устойчивое существование многих видов в экосистеме достигается за счет постоянно происходящих в ней естественных нарушений местообитаний, позволяющих новым поколениям занимать вновь освободившееся пространство.
Концепция экосистемы
Основным объектом изучения экологии являются экологические системы, или экосистемы. Экосистема занимает следующее после биоценоза место в системе уровней живой природы. Говоря о биоценозе, мы имели в виду только живые организмы. Если же рассматривать живые организмы (биоценоз) в совокупности с факторами окружающей среды, то это уже экосистема. Таким образом, экосистема — природный комплекс (биокосная система), образованный живыми организмами (биоценоз) и средой их обитания (например, атмосфера — косной, почва, водоем — биокосной и т.д.), связанными между собой обменом веществ и энергии.
Общепринятый в экологии термин «экосистема» ввел в 1935 г. английский ботаник А. Тенсли. Он считал, что экосистемы, «с точки зрения эколога представляют собой основные природные единицы на поверхности земли», в которые входит «не только комплекс организмов, но и весь комплекс физических факторов, образующих то, что мы называем средой биома, — факторы местообитания в самом широком смысле». Тенсли подчеркивал, что для экосистем характерен разного рода обмен веществ не только между организмами, но и между органическим и неорганическим веществом. Это не только комплекс живых организмов, но и сочетание физических факторов.
Экосистема (экологическая система) — основная функциональная единица экологии, представляющая собой единство живых организмов и среды их обитания, организованное потоками энергии и биологическим круговоротом веществ. Это фундаментальная общность живого и среды его обитания, любая совокупность совместно обитающих живых организмов и условий их существования (рис. 8).

Рис. 8. Различные экосистемы: а — пруда средней полосы (1 — фитопланктон; 2 — зоопланктон; 3 — жуки-плавунцы (личинки и взрослые особи); 4- молодые карпы; 5 — щуки; 6 — личинки хорономид (комаров-дергунцов); 7- бактерии; 8 — насекомые прибрежной растительности; б — луга (I — абиотические вещества, т.е. основные неорганические и органические слагаемые); II- продуценты (растительность); III- макроконсументы (животные): А — травоядные (кобылки, полевые мыши и т.д.); В — косвенные или питающиеся детритом консументы, или сапробы (почвенные беспозвоночные); С- «верховые» хищники (ястребы); IV- разлагатели (гнилостные бактерии и грибы)
Понятие «экосистема» можно применить к объектам различной степени сложности и величины. Примером экосистемы может служить тропический лес в определенном месте и в конкретный момент времени, населенный тысячами видов живущих вместе растений, животных и микробов и связанный происходящими между ними взаимодействиями. Экосистемами являются такие природные образования, как океан, море, озеро, луг, болото. Экосистемой может быть кочка на болоте и гниющее дерево в лесу с живущими на них и в них организмами, муравейник с муравьями. Самой большой экосистемой является планета Земля.
Каждая экосистема может характеризоваться определенными границами (экосистема елового леса, экосистема низинного болота). Однако само понятие «экосистема» безранговое. Она обладает признаком безразмерности, ей не свойственны территориальные ограничения. Обычно экосистемы разграничиваются элементами абиотической среды, например рельефом, видовым разнообразием, физико-химическими и трофическими условиями и т.н. Размер экосистем не может быть выражен в физических единицах измерения (площадь, длина, объем и т.д.). Он выражается системной мерой, учитывающей процессы обмена веществ и энергии. Поэтому под экосистемой обычно понимают совокупность компонентов биотической (живые организмы) и абиотической среды, при взаимодействии которых происходит более или менее полный биотический круговорот, в котором участвуют продуценты, консументы и редуценты. Термин «экосистема» применяется и по отношению к искусственным образованиям, например экосистема парка, сельскохозяйственная экосистема (агроэкосистема).
Экосистемы можно разделить на микроэкосистемы (дерево в лесу, прибрежные заросли водных растений), мезоэкосистемы (болото, сосновый лес, ржаное поле) и макроэкосистемы (океан, море, пустыня).
О равновесии в экосистемах
Равновесными называются такие экосистемы, которые «контролируют» концентрации биогенов, поддерживая их равновесие с твердыми фазами. Твердые же фазы (остатками живых организмов) являются продуктами жизнедеятельности биоты. Равновесными будут и те сообщества и популяции, которые входят в равновесную экосистему. Такой вид биологического равновесия называется подвижным , поскольку процессы отмирания непрерывно компенсируются появлением новых организмов.
Равновесные экосистемы подчиняются принципу устойчивости Лe Шателье. Следовательно, эти экосистемы обладают гомеоста- зом, — иными словами, способны минимизировать внешнее воздействие при сохранении внутреннего равновесия. Устойчивость экосистем достигается не смещением химических равновесий, а путем изменения скоростей синтеза и разложения биогенов.
Особый интерес представляет способ поддержания устойчивости экосистем, основанный на вовлечении в биологический круговорот органического веществ, ранее произведенного экосистемой и отложенного «про запас» — древесины и мортмассы (торф, гумус, подстилка). В этом случае древесина служит как бы индивидуальным материальным богатством, а мортмасса — коллективным, принадлежащим экосистеме в целом. Это «материальное богатство» увеличивает запас устойчивости экосистем, обеспечивая их выживание при неблагоприятных изменениях климата, стихийных бедствиях и др.
Устойчивость экосистемы тем больше, чем больше она по размеру и чем богаче и разнообразнее ее видовой и популяционный состав.
Экосистемы разного типа используют различные варианты индивидуальных и коллективных способов запасания устойчивости при различном соотношении индивидуального и коллективного материального богатства.
Таким образом, основная функция совокупности живых существ (сообщества), входящих в экосистему, — обеспечить равновесное (устойчивое) состояние экосистемы на основе замкнутого круговорота веществ.
Естественные экосистемы существуют сотни и тысячи лет, и обладают определенной стабильностью во времени и пространстве.
В естественной экосистеме постоянно поддерживается равновесие, исключающее необратимое уничтожение тех или иных звеньев трофической сети. Любая экосистема всегда сбалансирована и устойчива (гомеостатична), причем системы тем стабильнее во времени и пространстве, чем они сложнее.
Объект управления, существующий в экосистеме.| Экологическая система из двух популяций. В естественных экосистемах получение ресурсов и избавление от отходов происходит в рамках круговорота всех элементов.
В естественных экосистемах центральным звеном является растительность, и все процессы изменения в данных сообществах относятся к автотрофным сукцессиям.
В естественных экосистемах, не испытавших в значительной степени антропогенного воздействия, необходимое качество окружающей природной среды обеспечивается самой природой. При ведении хозяйственной деятельности в таких условиях задача состоит в том, чтобы не нарушать сложившегося баланса. В нарушенных экосистемах улучшение качества окружающей природной среды достигается регулированием степени удовлетворения экономических интересов предприятий - производителей и требований экологической защиты природы.
В естественных экосистемах гомеостаз поддерживается тем, что они открыты, т.е. постоянно обмениваются веществом и энергией с окружающей средой. В антропогенных, созданных под влиянием человека системах для поддержания гомеостаза необходимо вмешательство (управление) людей.
В естественных экосистемах, не испытавших в значительной степени антропогенного воздействия, качество окружающей природной среды обеспечивается самой природой. При ведении хозяйственной деятельности в таких условиях задача состоит в том, чтобы не нарушать сложившегося баланса. В нарушенных экосистемах улучшение качества окружающей природной среды достигается регулированием степени удовлетворения экономических интересов предприятий-производителей и требований экологической защиты природы.
В естественных экосистемах благоприятное качество среды обитания для доминирующих видов обеспечивается балансом энергии и вещества.
Ухудшение состояния естественных экосистем связано с нарушением в них многих природных процессов и ведет к утрате естественной способности к саморегуляции и воспроизводству природных экосистем. Загрязнение нефтепродуктами и промышленными стоками надолго сохраняется в среде, представлявшей основу для существования коренного населения Севера.
В почвах естественных экосистем эти процессы находятся в равновесии.
Наряду с естественными экосистемами существуют искусственные экосистемы, которые характеризуются значительно меньшим видовым разнообразием и обычно используются на ранних стадиях развития. Искусственные системы широко используются в сельском хозяйстве. Эти системы получают дополнительное количество энергии при обработке почвы в виде удобрений, пестицидов. Доминирующие виды растений и животных в агроэкосистемах получены путем искусственного, а не естественного отбора, как это имеет место в природных системах.
Как известно, естественные экосистемы находятся в состоянии динамического равновесия. Их эволюция происходит в сторону все большей устойчивости к возможным воздействиям. Мало того, определенные нагрузки могут увеличивать полезную продуктивность некоторых экосистем. Отсюда следует важный практический вывод о том, что не следует полностью воздерживаться от техногенного и иного воздействия на экосистемы из-за страха перед их неустойчивостью. Необходимо направлять усилия на тщательное изучение допустимых нагрузок на них. Обоснованное управление этими нагрузками является одним из условий устойчивого развития общества.
Каждый организм в естественной экосистеме производит потенциально загрязняющие среду отходы. Устойчивость экосистемы обусловлена тем, что отходы одних организмов становятся пищей и / или сырьем для других. В сбалансированных экосистемах отходы не накапливаются до уровня, вызывающего неблагоприятные изменения, а разлагаются и рециклизуются.
Поддержание замкнутых круговоротов в естественных экосистемах возможно благодаря наличию редуцентов, которые используют все отходы и остатки, и постоянному поступлению солнечной энергии. В городских и искусственных экосистемах редуценты отсутствуют или их количество ничтожно мало, поэтому наряду с другими причинами происходит накопление отходов, которые, накапливаясь, загрязняют окружающую среду. Для быстрейшего разложения и вторичного использования таких отходов следует создавать условия для развития редуцентов, например, путем компостирования. Так человек учится у природы.
Поддержание замкнутых круговоротов в естественных экосистемах возможно благодаря наличию разложителей (редуцентов), которые используют все отходы и остатки, и постоянному поступлению солнечной энергии. В городских и искусственных экосистемах мало или совсем нет редуцентов, и отходы (жидкие, твердые и газообразные) накапливаются, загрязняя окружающую среду. Способствовать быстрейшему разложению и вторичному использованию таких отходов можно, поощряя развитие редуцентов, например, путем компостирования. Так человек учится у природы.
Мутуализм), В естественных экосистемах преобладает ассоциативная А. В агроэкосистемах роль ассоциативной Б.а. резко снижается и не превышает 40 кг / га азота в год. По этой причине для активизации Б.а. возделывают бобовые растения. В средней полосе поле клевера или люцерны способно накопить за вегетационный сезон 200 - 400 кг / га азота, что полностью покрывает потребности в нем даже при интенсивном растениеводстве.
Правило внутренней непротиворечивости: в естественных экосистемах деятельность входящих в них видов направлена на поддержание этих экосистем как среды собственного обитания.
Правило внутренней непротиворечивости - в естественных экосистемах деятельность входящих в них видов направлена на поддержание этих экосистем как среды собственного обитания.
Примечательно, что растения в естественных экосистемах полностью зависят от собственных средств защиты против насекомых и других травоядных - лишнее доказательство того, насколько эффективными могут быть природные оборонительные средства. Многие из участвующих в этом химических соединений, в частности танины и алкалоиды, имеют горький вкус и многие токсичны для млекопитающих и других животных. Селекционные программы часто бывали направлены на снижение концентраций таких веществ в культурных растениях. В свете наших современных представлений о естественных химических средствах защиты не кажется странным то обстоятельство, что многие культурные растения сравнительно чувствительны к поеданию насекомыми. Поскольку многие культурные сорта довольно однородны в генетическом отношении, практически все особи данного сорта могут быть в равной степени чувствительны к нападениям насекомых. Очевидно, дело здесь в том, что селекция культурных растений, как правило, ведется с целью получения определенных структурных признаков, и эти изменения могут ослабить защитные механизмы растений против насекомых. Кроме того, большие группы сходных растений насекомым найти легче, чем обособленные особи, обычно встречающиеся в естественных экосистемах.
Экологическое неблагополучие - следствие прямого уничтожения естественных экосистем (вырубка лесов, распашка степей и лугов, осушение болот и пр.
Происходящее в настоящее время быстрое разрушение естественных экосистем, регулирующих окружающую среду, ведет к экологической катастрофе. Эта катастрофа, в свою очередь, сопровождается резким снижением темпов роста населения и стабилизацией его на уровне 7.39 млрд человек.
Многие потенциально патогенные бактерии являются компонентами естественных экосистем. На полях орошения выделяют иерсинии, цитробактеры, серрации, хафнии и др. Они проникают из почвы и воды в корневую систему растений и достигают высоких концентраций в их вегетативных органах. Эти бактерии тесно связаны с беспозвоночными в почве и воде - амебами, креветками, нематодами и др. Идет невидимая человеку битва. В ней находит применение и оттачивается весь арсенал факторов патогенности, который при подходящих условиях, связанных с изменением экологических характеристик внешней и внутренней среды, может быть использован против человека. Особенно опасны для сапрофитов простейшие. Разные виды простейших питаются различными видами микроорганизмов: кальпидиум и кальпида предпочитают некоторые виды псевдомонад; инфузория туфелька - дрожжи и псевдовульгарис. В свою очередь бактерии, защищаясь, вызывают целые эпизоотии среди простейших.
Практические наблюдения подтверждают, что в ненарушенных естественных экосистемах - действительно соблюдается такое условие.
Переход к устойчивому развитию предполагает постепенное восстановление естественных экосистем до уровня, гарантирующего стабильность окружающей среды. Этого можно достичь усилиями всего человечества, но начинать движение к данной цели каждая страна должна самостоятельно.
Переход к устойчивому развитию предполагает постепенное развитие естественных экосистем до уровня, гарантирующего стабильность окружающей среды. Этого можно достичь усилиями всего человечества, но начинать движение к поставленной цели каждая страна должна самостоятельно.
Интенсивная хозяйственная деятельность человека ведет к замене естественных экосистем искусственно созданными, в первую очередь сельскохозяйственными, или агроценозами.
Переход к устойчивому развитию предполагает постепенное восстановление естественных экосистем до уровня, гарантирующего стабильность окружающей среды, и должен обеспечить на перспективу сбалансированное решение проблем социально-экономического развития и сохранение благоприятной окружающей среды и природно-ресурсного потенциала.
Важнее правило относительной внутренней непротиворечивости: в естественных экосистемах (и прежде всего в их биотических компонентах, составляющих биоценозы) деятельность входящих в них видов направлена на поддержание этих систем как среды собственного обитания. Это правило было сформулировано автором в 50 - х гг. во время исследования роли позвоночных животных в жизни леса. Выше оно соотнесено с популяциями как правило сохранения видовой среды обитания (разд. Здесь мы обращаем внимание на то, что не только один вид (его популяция), но и их сообщество подчинено аналогичной закономерности.
Населению необходимо понять и знать, что разрушение естественных экосистем и сопутствующее исчезновение многих растений и животных становятся одним из основных факторов, ограничивающих экономический рост и жизнедеятельность человека па земле.
Вместе с тем в России сохранился крупнейший на планете массив естественных экосистем (8 млн. км2), который служит резервом устойчивости биосферы.
Но за время своего существования оно уже уничтожило 70 % естественных экосистем, которые способны переработать все отходы...
Третье ограничение относится преимущественно к интенсификации промысла, агротехническому преобразованию естественных экосистем и созданию хозяйственных инфраструктур.
В природе подсмотреть структуру такого кентавра с гелитроф-ным питанием, как у естественных экосистем и искусственной трофической структуры, которую формирует человек, невозможно - она получается расчетным путем в результате деятельности коллектива МДК Агроэкология, решающего задачи формулирования экологического императива и конструирования сестайнинга.
В целом, несмотря на многообразие форм проявления В.р., их роль в естественных экосистемах не следует переоценивать. Основным типом горизонтальных взаимоотношений растений является конкуренция.
Проблемными местами являются высокая стоимость проекта и необходимость государственного участия в его финансировании, а также нарушение естественных экосистем при добыче или транспортировке нефти и газа на территории Национальных парков.
На определяющее значение в круговороте веществ этих групп организмов указывает также баланс потребления чистой первичной продукции в естественных экосистемах.
Следует отметить, что влияние загрязняющих веществ не имеет видимых границ и барьеров, оно способно действовать на естественные экосистемы, находящиеся преимущественно в состоянии природного равновесия. Поскольку химическое загрязнение - относительно недавнее явление, на таких практически фоновых территориях оно характеризуется лишь первыми начальными стадиями.
Для обеспечения экологической безопасности человечества возможны три направления его деятельности: 1) сохранение и поддержание расширенного воспроизводства естественных экосистем; 2) управление природно-хозяйственньши системами, средообразую-щие функции которых близки к естественным экосистемам, место которых они заняли; 3) реализация экологически чистых технологий, устраняющих последствия хозяйственной деятельности, неблагоприятные для биосферы и человека.
Типы динамики населения по С А. Север-цову. Характер закономерных изменений численности видоспецифичен и в целом связан с особенностями биологии вида, его физиологии и места в естественных экосистемах. Еще в начале 40 - х годов нашего века С.А. Северцов, проанализировавший многолетний ход численности у большого числа видов млекопитающих и птиц, установил несколько типов ее динамики.
Таким образом, возникновение зон чрезвычайной экологической ситуации характеризуется определенными изменениями окружающей среды, состояния здоровья человека, а также деградацией естественных экосистем.
Вследствие зависимости растительности от реакции почвы (Boeker, 1964) при подкислении могут происходить изменения видового разнообразия сенокосных угодий и естественных экосистем. Для сельскохозяйственных площадей важно, чтобы в почве имелось достаточное количество кальция, необходимого для поддержания рН в оптимальных пределах, обеспечивающих протекание реакций обмена ионов, насыщения коллоидов и поддержания рыхлости почвы. В качестве питательного вещества кальций, кроме того, повышает устойчивость к SO2 и HF (см. разд.
В реальности, если учесть самоочищающий потенциал северных рек, приведенные цифры в 10 - 12 раз занижены по отношению к естественным экосистемам.
Второй принцип: природные ресурсы Земли, включая воздух, воду, землю, флору, фауну и особенно репрезентативные (характерные) образцы естественных экосистем, должны быть сохранены на благо нынешнего и будущих поколений путем тщательного планирования деятельности человека и управления ею по мере необходимости.
Во-вторых, стратегия реализации концепции перехода России к устойчивому развитию предполагает активное проведение в жизнь мер и действий по восстановлению качества окружающей среды и естественных экосистем в промышленных регионах, густонаселенных и других районах, с повышенной технологией и экологической опасностью.
Понятие экосистемы
Определения
Иногда особо подчёркивается, что экосистема - это исторически сложившаяся система (см. Биоценоз).
Концепция экосистемы

Юджин Одум (1913-2000). Отец экосистемной экологии
Экосистема - сложная (по определению сложных систем Л. Берталанфи) самоорганизующаяся, саморегулирующаяся и саморазвивающаяся система. Основной характеристикой экосистемы является наличие относительно замкнутых, стабильных в пространстве и времени потоков вещества и энергии между биотической и абиотической частями экосистемы . Из этого следует, что не всякая биологическая система может назваться экосистемой, например, Таковыми не являются аквариум или трухлявый пень . Данные биологические системы (естественные или искусственные) не являются в достаточной степени самодостаточными и саморегулируемыми (аквариум), если перестать регулировать условия и поддерживать характеристики на одном уровне, достаточно быстро она разрушится. Такие сообщества не формируют самостоятельных замкнутых циклов вещества и энергии (пень), а являются лишь частью большей системы . Такие системы следует называть сообществами более низкого ранга, или же микрокосмами . Иногда для них употребляют понятие - фация (например, в геоэкологии), но оно не способно в полной мере описать такие системы, особенно искусственного происхождения. В общем случае в разных науках понятию «фация» соответствуют различные определения: от систем субэкосистемного уровня (в ботанике, ландшафтоведении) до понятий, не связанных с экосистемой (в геологии), либо понятие, объединяющее однородные экосистемы (Сочава В. Б.), или почти тождественное (Берг Л. С. , Раменский Л. Г.) определению экосистемы.
Биогеоценоз и экосистема
В соответствии с определениями между понятиями «экосистема» и «биогеоценоз» нет никакой разницы, биогеоценоз можно считать полным синонимом термина экосистема . Однако существует распространённое мнение, согласно которому биогеоценоз может служить аналогом экосистемы на самом начальном уровне , так как термин «биогеоценоз» делает бо́льший акцент на связь биоценоза с конкретным участком суши или водной среды, в то время как экосистема предполагает любой абстрактный участок. Поэтому биогеоценозы обычно считаются частным случаем экосистемы . Разными авторами в определении термина биогеоценоз перечисляются конкретные биотические и абиотические компоненты биогеоценоза, в то время как определение экосистемы носит более общий характер .
Строение экосистемы
В экосистеме можно выделить два компонента - биотический и абиотический. Биотический делится на автотрофный (организмы, получающие первичную энергию для существования из фото- и хемосинтеза или продуценты) и гетеротрофный (организмы, получающие энергию из процессов окисления органического вещества - консументы и редуценты) компоненты , формирующие трофическую структуру экосистемы.
Единственным источником энергии для существования экосистемы и поддержания в ней различных процессов являются продуценты, усваивающие энергию солнца , (тепла , химических связей) с эффективностью 0,1 - 1 %, редко 3 - 4,5 % от первоначального количества. Автотрофы представляют первый трофический уровень экосистемы. Последующие трофические уровни экосистемы формируются за счёт консументов (2-ой, 3-й, 4-й и последующие уровни) и замыкаются редуцентами , которые переводят неживое органическое вещество в минеральную форму (абиотический компонент), которая может быть усвоена автотрофным элементом .
Основные компоненты экосистемы
С точки зрения структуры в экосистеме выделяют :
- климатический режим, определяющий температуру, влажность, режим освещения и прочие физические характеристики среды;
- неорганические вещества, включающиеся в круговорот;
- органические соединения, которые связывают биотическую и абиотическую части в круговороте вещества и энергии;
- продуценты - организмы, создающие первичную продукцию;
- макроконсументы, или фаготрофы, - гетеротрофы, поедающие другие организмы или крупные частицы органического вещества;
- микроконсументы (сапротрофы) - гетеротрофы, в основном грибы и бактерии , которые разрушают мёртвое органическое вещество, минерализуя его, тем самым возвращая в круговорот.
Последние три компонента формируют биомассу экосистемы.
С точки зрения функционирования экосистемы выделяют следующие функциональные блоки организмов (помимо автотрофов):
- биофаги - организмы, поедающие других живых организмов,
- сапрофаги - организмы, поедающие мёртвое органическое вещество.
Данное разделение показывает временно-функциональную связь в экосистеме, фокусируясь на разделении во времени образования органического вещества и перераспределении его внутри экосистемы (биофаги) и переработки сапрофагами . Между отмиранием органического вещества и повторным включением его составляющих в круговорот вещества в экосистеме может пройти существенный промежуток времени, например, в случае соснового бревна, 100 и более лет.
Все эти компоненты взаимосвязаны в пространстве и времени и образуют единую структурно-функциональную систему.
Экотоп
Обычно понятие экотоп определялось как местообитание организмов, характеризующееся определённым сочетанием экологических условий: почв, грунтов, микроклимата и др . Однако, в этом случае это понятие фактически почти идентично понятию климатоп .
На данный момент под экотопом в отличие от биотопа понимается определённая территория или акватория со всем набором и особенностями почв , грунтов , микроклимата и других факторов в неизменённом организмами виде . Примерами экотопа могут служить наносные грунты, новообразовавшиеся вулканические или коралловые острова , вырытые человеком карьеры и другие заново образовавшиеся территории. В этом случае климатоп является частью экотопа.
Климатоп
Изначально «климатоп» был определён В. Н. Сукачёвым (1964) как воздушная часть биогеоценоза, отличающаяся от окружающей атмосферы своим газовым составом, особенно концентарией углекислого газа в приземном биогоризонте, кислорода там же и в биогоризонтах фотосинтеза , воздушным режимом, насыщенностью биолинами, уменьшенной и изменённой солнечной радиацией и освещённостью, наличием люминесценции растений и некоторых животных, особым тепловым режимом и режимом влажности воздуха .
На данный момент это понятие трактуется чуть более широко: как характеристика биогеоценоза, сочетание физических и химических характеристик воздушной или водной среды, существенных для населяющих эту среду организмов . Климатоп задаёт в долговременном масштабе основные физические характеристики существования животных и растений, определяя круг организмов, которые могут существовать в данной экосистеме.
Эдафотоп
Под эдафотопом обычно понимается почва как составной элемент экотопа . Однако более точно это понятие следует определять как часть косной среды преобразованной организмами, то есть не всю почву , а лишь её часть . Почва (эдафотоп) является важнейшей составляющей экосистемы: в нём происходит замыкание циклов вещества и энергии, осуществляется перевод из мёртвого органического вещества в минеральные и их вовлечение в живую биомассу . Основными носителями энергии в эдафотопе выступают органические соединения углерода , их лабильные и стабильные формы, они в наибольшей степени определяют плодородие почв.

Биоценоз, представленный в схематичном виде как пищевая сеть и его биотоп
Биотоп
Биоценоз
Иногда выделяют третий аспект устойчивости - устойчивость экосистемы по отношению к изменениям характеристик среды и изменению своих внутренних характеристик . В случае, если экосистема устойчиво функционирует в широком диапазоне параметров окружающей среды и/или в экосистеме присутствует большое число взаимозаменяемых видов (то есть, когда различные виды, сходные по экологическим функциям в экосистеме, могут замещать друг друга), такое сообщество называют динамически прочным (устойчивым). В обратном случае, когда экосистема может существовать в весьма ограниченном наборе параметров окружающей среды, и/или большинство видов незаменимы в своих функциях, такое сообщество называется динамически хрупким (неустойчивым) . Необходимо отметить, что данная характеристика в общем случае не зависит от числа видов и сложности сообществ. Классическим примером может служить Большой Барьерный риф у берегов Австралии (северо-восточное побережье), являющийся одной из «горячих точек» биоразнообразия в мире - симбиотические водоросли кораллов, динофлагелляты , весьма чувствительны к температуре. Отклонение от оптимума буквально на пару градусов ведёт к гибели водорослей, а до 50-60 % (по некоторым источникам до 90 %) питательных веществ полипы получают от фотосинтеза своих мутуалистов .
У экосистем существует множество состояний, в которых она находится в динамическом равновесии; в случае выведения из него внешними силами, экосистема совершенно необязательно вернётся в изначальное состояние, зачастую её привлечёт ближайшее равновесное состояние (аттрактор), хотя оно может быть очень близким к первоначальному .
Биоразнообразие и устойчивость в экосистемах

Дождевые леса Амазонии, как и влажные экваториальные леса, являются местами наибольшего биоразнообразия
Обычно устойчивость связывали и связывают с биоразнообразием видов в экосистеме (альфаразнообразие), то есть, чем выше биоразнообразие, чем сложнее организация сообществ, чем сложнее пищевые сети, тем выше устойчивость экосистем. Но уже 40 и более лет назад на данный вопрос существовали различные точки зрения, и на данный момент наиболее распространено мнение, что как локальная, так и общая устойчивость экосистемы зависят от значительно большего набора факторов, чем просто сложность сообществ и биоразнообразие. Так, на данный момент с повышением биоразнообразия обычно связывают повышение сложности, силы связей между компонентами экосистемы, стабильность потоков вещества и энергии между компонентами .

Экваториальный дождевой лес может содержать более 5000 видов растений (для сравнения в лесах таёжной зоны - редко более 200 видов)
Важность биоразнообразия состоит в том, что оно позволяет формировать множество сообществ, различных по структуре, форме, функциям, и обеспечивает устойчивую возможность их формирования. Чем выше биоразнообразие, тем большее число сообществ может существовать, тем большее число разнообразных реакций (с точки зрения биогеохимии) может осуществляться, обеспечивая существование биосферы в целом .
Сложность и устойчивость экосистем
На данный момент не существует удовлетворительного определения и модели, описывающей сложность систем и экосистем в частности. Существует два широко распространённых определения сложности: колмогоровская сложность - слишком специализированное для применения к экосистемам. И более абстрактное, но тоже неудовлетворительное определение сложности, данное И. Пригожиным в работе «Время, хаос, квант» : Сложные системы - не допускающие грубого или операционального описания в терминах детерминистских причинностей . В других своих трудах И. Пригожин писал, что не готов дать строгого определения сложности , поскольку сложное - это нечто, что на данный момент не может быть корректно определено.
Параметры сложности и их влияние на устойчивость
В качестве параметров сложности экосистем традиционно подразумевались общее число видов (альфа-разнообразие), большое число взаимодействий между видами, сила взаимодействий между популяциями и различные сочетания этих характеристик. При дальнейшем развитии этих представлений появилось утверждение, что чем больше путей переноса и преобразования энергии в экосистеме, тем она устойчивей при различных видах нарушений .
Однако, позже было показано, что данные представления не могут охарактеризовать устойчивость экосистем . Существует множество примеров как весьма устойчивых монокультурных сообществ (фитоценозы орляка), так и слабоустойчивых сообществ с высоким биоразнообразием (коралловые рифы, тропические леса). В 70-80-х годах XX столетия усилился интерес к моделированию зависимости устойчивости от сложности экосистем . Разработанные в этот период модели показали, что в случайным образом генерируемой сети взаимодействия в сообществе при удалении бессмысленных цепей (типа А ест В, В ест С, С ест А и подобного типа) локальная устойчивость падает с увеличением сложности. Если продолжить усложнение модели и учесть, что консументы испытывают влияние пищевых ресурсов, а пищевые ресурсы от консументов не зависят, то можно прийти к выводу о том, что устойчивость не зависит от сложности, либо также падает с её увеличением. Разумеется, такие результаты справедливы главным образом для детритных цепей питания, в которых консументы не влияют на поток пищевых ресурсов , хотя и могут менять пищевую ценность последних.
При изучении общей устойчивости на модели из 6 видов (2 хищника-консумента второго порядка, 2 консумента первого порядка и 2 вида в основании пищевой цепи) исследовалось удаление одного из видов. В качестве параметра устойчивости была принята связность. Сообщество считалось устойчивым, если остальные виды оставались локально устойчивыми. Полученные результаты согласовывались с общепринятыми воззрениями о том, что с повышением сложности при выпадении хищников высшего порядка устойчивость сообщества падает, но при выпадении оснований пищевой цепи с повышением сложности устойчивость повышалась .
В случае упругой устойчивости , когда под сложностью также понимается связность, с повышением сложности упругая устойчивость также повышается. То есть, большее разнообразие видов и большая сила связи между ними позволяет сообществам быстрее восстанавливать свою структуру и функции. Данный факт подтверждает общепринятые взгляды на роль биоразнообразия как некоего пула (фонда) для восстановления полноценной структуры как экосистем, так и более высокоорганизованных структур биосферы, а также самой биосферы в целом. На данный момент общепринятым и фактически неоспариваемым является представление о том, что биосфера эволюционировала в сторону увеличения биоразнообразия (всех трёх его компонентов), ускорения обращения вещества между компонентами биосферы, и «убыстрения» времени жизни как видов, так и экосистем .
Потоки вещества и энергии в экосистемах
На данный момент научное понимание всех процессов внутри экосистемы далеко от совершенства, и в большей части исследований либо вся экосистема, либо некоторые её части выступают в качестве «чёрного ящика » . В то же время, как любая относительно замкнутая система, экосистема характеризуется входящим и выходящим потоком энергии и распределением этих потоков между компонентами экосистем.
Продуктивность экосистем
При анализе продуктивности и потоков вещества и энергии в экосистемах выделяют понятия биомасса и урожай на корню . Под урожаем на корню понимается масса тел всех организмов на единице площади суши или воды , а под биомассой - масса этих же организмов в пересчёте на энергию (например, в джоулях) или в пересчёте на сухое органическое вещество (например, в тоннах на гектар) . К биомассе относят тела организмов целиком, включая и витализированные омертвевшие части и не только у растений, к примеру, кора и ксилема , но и ногти и ороговевшие части у животных. Биомасса превращается в некромассу только тогда, когда отмирает часть организма (отделяется от него) или весь организм. Часто зафиксированные в биомассе вещества являются «мёртвым капиталом», особенно это выражено у растений: вещества ксилемы могут сотнями лет не поступать в круговорот, служа только опорой растения .
Под первичной продукцией сообщества (или первичной биологической продукцией) понимается образование биомассы (более точно - синтез пластических веществ) продуцентами без исключения энергии, затраченной на дыхание за единицу времени на единицу площади (например, в сутки на гектар).
Первичную продукцию сообщества разделяют на валовую первичную продукцию , то есть всю продукцию фотосинтеза без затрат на дыхание , и чистую первичную продукцию , являющуюся разницей между валовой первичной продукцией и затратами на дыхание. Иногда её ещё называют чистой ассимиляцией или наблюдаемым фотосинтезом ).
Чистая продуктивность сообщества - скорость накопления органического вещества, не потребляемого гетеротрофами (а затем и редуцентами). Обычно вычисляется за вегетационный период либо за год . Таким образом, это часть продукции, которая не может быть переработана самой экосистемой. В более зрелых экосистемах значение чистой продуктивости сообщества стремится к нулю (см. концепцию климаксных сообществ).
Вторичная продуктивность сообщества - скорость накопления энергии на уровне консументов. Вторичную продукцию не подразделяют на валовую и чистую, так как консументы только потребляют энергию, усвоенную продуцентами, часть её не ассимилируется, часть идёт на дыхание, а остаток идёт в биомассу, поэтому более корректно называть её вторичной ассимиляцией .
Распределение энергии и вещества в экосистеме может быть представлено в виде системы уравнений. Если продукцию продуцентов представить как P 1 , то продукция консументов первого порядка будет выглядеть следующим образом:
- P 2 =P 1 -R 2 ,
где R 2 - затраты на дыхание, теплоотдача и неассимилированная энергия. Следующие консументы (второго порядка) переработают биомассу консументов первого порядка в соответствии с:
- P 3 =P 2 -R 3
и так далее, до консументов самого высшего порядка и редуцентов. Таким образом, чем больше в экосистеме потребителей (консументов), тем более полно перерабатывается энергия, первоначально зафиксированная продуцентами в пластических веществах . В климаксных сообществах, где разнообразие для данного региона обычно максимально, такая схема переработки энергии позволяет сообществам устойчиво функционировать на протяжении длительного времени.
Энергетические соотношения в экосистемах (экологические эффективности)

График изменения соотношения P/B в экосистемах (по А. К. Бродскому, 2002)
Пространственные границы экосистемы (хорологический аспект)
В природе, как правило, не существует чётких границ между различными экосистемами . Всегда можно указать на ту или иную экосистему, но выделить дискретные границы, если они не представлены различными ландшафтными факторами (обрывы, реки, различные склоны холмов, выходы скальных пород и т. п.), не представляется возможным, ибо чаще всего существуют плавные переходы от одной экосистемы к другой . Это обусловлено относительно плавным изменением градиента факторов среды (влажность, температура, увлажнённость и прочее). Иногда переходы из одной экосистемы в другую могут фактически являться самостоятельной экосистемой. Обычно сообщества, образующиеся на стыке различных экосистем, называются экотонами . Термин «экотон» введён Ф. Клементсом в 1905 году.
Экотоны
Экотоны играют существенную роль в поддержании биологического разнообразия экосистем за счёт так называемого краевого эффекта - сочетания комплекса факторов среды различных экосистем, обуславливающее большее разнообразие условий среды, следовательно, лицензий и экологических ниш . Тем самым возможно существование видов как из одной, так и из другой экосистемы, а также специфичных для экотона видов (например растительность прибрежно-водных местообитаний).

Некоторые возможные варианты границ (экотоны) между экосистемами
В российской литературе краевой эффект иногда называют эффектом опушки .
Примерами экотонов могут служить прибрежные участки суши и водоемов (например, литораль), опушки, переходы из лесных экосистем в полевые, эстуарии . Однако не всегда экотон является местом повышенного биоразнообразия видов. К примеру, эстуарии рек, впадающих в моря и океаны , наоборот, характеризуются пониженным биоразнообразием видов, так как средняя солёность дельт не позволяет существовать многим пресноводным и солоноводным (морским) видам.
Альтернативным представлением о континуальных переходах между экосистемами является представление о экоклинах (экологических рядах). Экоклин - постепенная смена биотопов, генетически и фенотипически приспособленных к конкретной среде обитания, при пространственном изменении какого-либо фактора среды (обычно климатического), а потому составляющих непрерывный ряд форм без заметных перерывов постепенности. Экоклин невозможно разделить на экотипы. Например, длина ушей лисиц и мн. др., их признаки изменяются с севера на юг настолько постепенно, что очень затруднительно выделить четкие морфологические группы, которые бы естественно объединялись в подвиды.
Временные границы экосистемы (хронологический аспект)

Смена сообщества в сосновом лесу после низового пожара (слева) и через два года после пожара (справа)
На одном и том же биотопе с течением времени существуют различные экосистемы. Смена одной экосистемы на другую может занимать как довольно длительные, так относительно короткие (несколько лет) промежутки времени. Длительность существования экосистем в таком случае определяется этапом сукцессии . Смена экосистем в биотопе может быть обусловлена и катастрофическими процессами, но в таком случае, существенно изменяется и сам биотоп, и такую смену не принято называть сукцессией (за некоторыми исключениями, когда катастрофа, например, пожар - естественный этап циклической сукцессии) .
Сукцессия
Сукцессия - это последовательная, закономерная смена одних сообществ другими на определённом участке территории, обусловленная внутренними факторами развития экосистем . Каждое предыдущее сообщество предопределяет условия существования следующего и собственного исчезновения . Это связано с тем, что в экосистемах, которые являются переходными в сукцессионом ряду, происходит накопление вещества и энергии, которые они уже не в состоянии включить в круговорот, преобразование биотопа, изменение микроклимата и других факторов, и тем самым создаётся вещественно-энергетическая база, а также и условия среды, необходимые для формирования последующих сообществ. Однако, есть и другая модель, которая объясняет механизм сукцессии следующим образом : виды каждого предыдущего сообщества вытесняются лишь последовательной конкуренцией, ингибируя и «сопротивляясь» внедрению последующих видов. Тем не менее, эта теория рассматривает лишь конкурентные отношения между видами, не описывая всю картину экосистемы в целом. Безусловно, такие процессы идут, но конкурентное вытеснение предыдущих видов возможно именно из-за преобразования ими биотопа. Таким образом, обе модели описывают разные аспекты процесса и верны одновременно.
Сукцессия бывает автотрофной (например, сукцессия после лесного пожара) и гетеротрофной (например, осушенное болото) . На ранних стадиях автотрофной сукцессионной последовательности соотношение P/R много больше единицы, так как обычно первичные сообщества обладают высокой продуктивностью, но структура экосистемы ещё не сформировалась полностью, и нет возможности утилизировать эту биомассу . Последовательно, с усложнением сообществ, с усложнением структуры экосистемы, расходы на дыхание (R) растут, так как появляется всё больше гетеротрофов, ответственных за перераспределение вещественно-энергетических потоков, соотношение P/R стремится к единице и фактически является таковым у терминального сообщества (экосистемы) . Гетеротрофная сукцессия обладает обратными характеристиками: в ней соотношение P/R на ранних этапах много меньше единицы (так как существует много органического вещества и нет необходимости в его синтезе, его можно сразу использовать на построение сообщества) и постепенно увеличивается по мере продвижения по сукцессионным стадиям.

Пример стадии гетеротрофной сукцессии - заболоченный луг
На ранних этапах сукцессии видовое разнообразие мало, но по мере развития разнообразие нарастает, изменяется видовой состав сообщества, начинают преобладать виды со сложными и продолжительными жизненными циклами, обычно появляются всё более крупные организмы, происходит развитие взаимовыгодных коопераций и симбиозов , усложняется трофическая структура экосистемы. Обычно предполагается, что терминальная стадия сукцессии обладает наибольшим видовым биоразнообразием. Это справедливо не всегда, но для климаксных сообществ тропических лесов это утверждение справедливо , а для сообществ умеренных широт пик разнообразия приходится на середину сукцессинного ряда или ближе к терминальной стадии . На ранних стадиях сообщества состоят из видов с относительно высокой скоростью размножения и роста, но низкой способностью к индивидуальному выживанию (r-стратеги). В терминальной стадии воздействие естественного отбора благоприятствует видам с низкой скоростью роста, но большей способностью к выживанию (k-стратеги).
По мере продвижения по сукцессионному ряду происходит всё большее вовлечение биогенных элементов в круговорот в экосистемах, возможно относительное замыкание внутри экосистемы потоков таких биогенных элементов, как азот и кальций (одни из наиболее подвижных биогенов) . Поэтому в терминальной стадии, когда большая часть биогенов вовлечена в круговорот, экосистемы более независимы от внешнего поступления данных элементов .
Для исследования процесса сукцессии применяют различные математические модели, в том числе стохастического характера
Климаксное сообщество
Понятие сукцессии тесно тесно связано с понятием климаксного сообщества . Климаксное сообщество формируется в результате последовательной смены экосистем и представляет собой наиболее сбалансированное сообщество , максимально эффективно использующее вещественно-энергетические потоки, то есть поддерживающее максимально возможную биомассу на единицу поступающей в экосистему энергии.
Теоретически у каждого сукцессионного ряда существует климаксное сообщество (экосистема), которое является терминальной стадией развития (или несколько, так называемая концепция поликлимакса). Однако, в реальности сукцессинный ряд замыкается климаксом не всегда, может реализоваться субклимаксное сообщество (или названное Ф. Клементсом - плагиклимакс), которое представляет собой сообщество, предшествующее климаксному, достаточно развитое структурно и функционально . Такая ситуация может возникать в силу естественных причин - условий среды или вследствие деятельности человека (в таком случае его называют дисклимакс ).
Ранги экосистем
Вопрос ранжирования экосистем достаточно сложен. Выделение минимальных экосистем (биогеоценозов) и экосистемы наивысшего ранга - биосферы не вызывает сомнений . Промежуточные же выделения довольно сложны, так как сложности хорологического аспекта не всегда однозначно позволяют определить границы экосистем. В геоэкологии (и ландшафтоведении) существует следующее ранжирование: фация - урочище (экосистема) - ландшафт - географический район - географическая область - биом - биосфера . В экологии существует сходное ранжирование , однако, обычно считается, что корректно выделение лишь одной промежуточной экосистемы - биома.
Биомы
Биом - крупное системно-географическое (экосистемное) подразделение в пределах природно-климатической зоны (Реймерс Н. Ф.). Согласно Р. Х. Уиттекеру - группа экосистем данного континента, которые имеют сходную структуру или физиономию растительности и общий характер условий среды. Это определение несколько некорректно, так как существует привязка к конкретному континенту, а некоторые биомы присутствуют на разных континентах, например, тундровый биом или степной .
На данный момент наиболее общепринятое определение звучит так: «Биом - совокупность экосистем со сходным типом растительности, расположенных в одной природно-климатической зоне» (Акимова Т. А., Хаскин В. В. ).
Общим в этих определениях является то, что в любом случае биомом называется совокупность экосистем одной природно-климатической зоны.
Выделяют от 8 до 30 биомов. Географическое распределение биомов определяют :
- Закон географической зональности (сформулированный В. В. Докучаевым)

| Наземные биомы, классифицированные по типу растительности | ||||
| Полярные пустыни Тундра Тайга Широколиственные леса | Степи Субтропические дождевые леса Средиземноморские биомы Муссонные леса | Аридные пустыни Ксерофитных кустарников Южные степи Семиаридные пустыни | Саванны Саванны с древесной растительностью (лесостепи) Субтропический лес Тропический дождевой лес | Альпийская тундра Горные леса |
Биосфера
Термин биосфера был введён Жаном-Батистом Ламарком в начале XIX века, а в геологии предложен австрийским геологом Эдуардом Зюссом в 1875 году . Однако создание целостного учения о биосфере принадлежит русскому учёному Владимиру Ивановичу Вернадскому .
Биосфера - экосистема высшего порядка, объединяющая все остальные экосистемы и обеспечивающая существование жизни на Земле. В состав биосферы входят следующие «сферы» :
Биосфера тоже не замкнутая система, она фактически полностью обеспечивается энергией Солнца , небольшую часть составляет тепло самой Земли. Ежегодно Земля получает от Солнца около 1,3*10 24 калорий . 40 % от этой энергии излучается обратно в космос, около 15 % идёт на нагрев атмосферы, почвы и воды, вся остальная энергия является видимым светом, который и является источником фотосинтеза.
В. И. Вернадский впервые чётко сформулировал понимание того, что всё живое на планете неразрывно связанно с биосферой и обязано ей своим существованием:
Искусственные экосистемы
Искусственные экосистемы - это экосистемы, созданные человеком, например, агроценозы , природно-хозяйственные системы или Биосфера 2 .
Искусственные экосистемы имеют тот же набор компонентов, что и естественные: продуценты , консументы и редуценты , но есть существенные отличия в перераспределении потоков вещества и энергии. В частности, созданные человеком экосистемы отличаются от естественных следующим :
Без поддержания энергетических потоков со стороны человека в искусственных системах с той или иной скоростью восстанавливаются естественные процессы и формируется естественная структура компонентов экосистемы и вещественно-энергетических потоков между ними.
Понятия, сходные с понятием экосистемы, в смежных науках
В экогеологии, ландшафтоведении и геоэкологии
В данных науках существуют понятия, схожие с понятием экосистема. Различие состоит в том, что в данных науках происходит смещение аспекта рассмотрения структуры и функций экосистем.
В целом, в географических науках принято рассматривать природный территориальный комплекс , как эквивалент экосистемы.
См. также
Примечания
- Forbes, S. A. The lake as a microcosm (англ.) // Bull. Sci. Assoc . - Peoria, Illinois, 1887. - P. 77–87. Reprinted in Illinois Nat. Hist. Survey Bulletin 15(9):537-550.
- Ю. Одум. Основы экологии. - М .: Мир, 1975. - 741 с.
- . Словари на Академке. Архивировано
- Ю. Одум. Экология. - М .: Мир, 1986.
- Раздел «Экосистемы» . The ECOLOGY site. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Биогеоценоз Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Николайкин, Н. И. , Николайкина, Н. Е., Мелехова, О. П. Экология. - 5-е. - М.: Дрофа, 2006. - 640 с.
- Бродский А. К. Краткий курс общей экологии, Учебное пособие для ВУЗов. - СПб: «Деан», 2000. - 224 с.
- Н. В. Короновский , Гидротермальные образования в океанах . Соросовский Образовательный Журнал, - №10, 1999, - cтр.55-62. Проверено 14 августа 2010.
- Д. В. Гричук. Теродинамические модели субмаринных гидротермальных систем . - М .: Научный мир, 2000. - ISBN УДК 550.40
- В. Ф. Левченко. Глава 3 // Эволюция Биосферы до и после появления человека. - СПб: Наука, 2004. - 166 с. - ISBN 5-02-026214-5
- Раутиан А. С. Палеонтология как источник сведений о закономерностях и факторах эволюции // Современная палеонтология . - М ., 1988. - Т. 2. - С. 76-118.
- Раутиан А. С., Жерихин В. В. Модели филоценогенеза и уроки экологических кризисов геологического прошлого // Журн. общ. биологии . - 1997. - Т. 58 № 4. - С. 20-47.
- Остроумов С. А. Новые варианты определений понятий и терминов «экосистема» и «биогеоценоз» // ДАН . - 2002. - Т. 383 № 4. - С. 571-573.
- М. Бигон, Дж. Харпер, К. Таунсенд. Экология. Особи, популяции и сообщества. - М .: Мир, 1989.
- Экотоп Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Т. А. Работнов «О Биогеоценозах» . // Бюллетень МОИП, отдел биологический, - т. 81, - вып. 2. - 1976. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Климатоп . Быков Б. А. "Экологический словарь" - Алма-Ата: "Наука", 1983 - с.216. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Основные термины экологии . Буренина Е.М., Буренин Е.П. Электронный учебник по экологии.. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Климатоп . Словарь по естественным наукам (Яндекс словари). Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Эдафотоп Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- . Экологический словарь(Словари на Академике). Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Биоценоз . Большая Советская Энциклопедия. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Зооценоз . Большая Советская Энциклопедия. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Гомеостаз экосистемы . Научно-Информационный портал ВИНИТИ. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Хатчинсон С., Хоукинс Л. Е. Океаны. Энциклопедический путеводитель. - М .: Махаон, 2007. - 304 с. - ISBN 5-18-001089-6
- А. Гиляров. «Кораллы обесцвечиваются из-за утраты взаимопонимания» . Элементы большой науки. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- А. Д. Арманд , Эксперимент «Гея», проблема живой земли . Российская Академия Наук. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- А. В. Галанин. Лекции по экологии. . Сайт Ботанического сада ДВО РАН. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Пригожин И., Стенгерс И. Время, хаос, квант . - Москва, 1994. - С. 81. - 263 с.
- Николис Г., Пригожин И. Познание сложного. - М.: Мир, 1990. - с. 352. Страница 47
- MacArthur R.H. Fluctuations of animal populations and a measure of community stability // Ecology, 36, 1955, - pp. 533-536
- May R.M. Will a large complex system be stable? // Nature (London), 1972, 238, - pp. 413-414
- May R.M Models for single populations. // Theoretical Ecology: Principles and Applications, 2nd edn., R.M. May ed. - pp. 5-29, - Blackwell Scientific Publications, Oxford 1981
- May R.M Models for two interacting populations. // Theoretical Ecology: Principles and Applications, 2nd edn., R.M. May ed. - pp.78-104, - Blackwell Scientific Publications, Oxford 1981
- May R.M Patterns in multi-species communities. // Theoretical Ecology: Principles and Applications, 2nd edn., R.M. May ed., - Blackwell Scientific Publications, Oxford 1981
- DeAngelis D.L. Stability and connectance in food web models // Ecology 56, 1975, - pp. 238-243
- Pimm S.L. The structure of food webs // Theoretical Population Biology, 16, 1979, - pp. 144-158
- Pimm S.L. Complexity and stability: another look at MacArthu’s original hypothesis // Oikos , 33, 1979, - pp. 351-357
- В. Ф. Левченко, Я. И. Старобогатов Физико-экологический подход к эволюции биосферы . // «Эволюционная биология: история и теория». Санкт-Петербург, 1999, - с. 37-46. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Левченко В. Ф. Эволюция биосферы до и после появления человека. . Санкт-Петербург, Институт эволюционной физиологии и биохимии Российской Академии наук, - Издательство «НАУКА», 2004. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Первичная продукция . Научно-информационный портал ВИНИТИ. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Первичная продуктивность . Глоссарий.ру. Проверено 14 августа 2010.
- Маврищев В.В. Континуум, экотоны, краевой эффект // Основы экологии: учебник . - 3-е изд. испр. и доп. - Минск: Высшая школа, 2007. - 447 с. - ISBN 978-985-06-1413-1
- Экотон. . Словарь по естественным наукам (Яндекс словари). Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Экотон и концепция краевого (пограничного) эффекта . сайт Биоэкология. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Кревой эффект . Экологический энциклопедический словарь. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Эстуарий. . Словарь терминов по физической географии Института географии РАН. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Сукцессия . Большая Советская Энциклопедия. Архивировано из первоисточника 22 августа 2011. Проверено 14 августа 2010.
- Развитие и эволюция экосистемы . портал Инженерная экология.