Влияние симпатических нервов на деятельность сердца. Симпатическая и парасимпатическая нервная система
Читайте также
Вегетативная нервная система (ВНС) – отдел нервной системы, регулирующий деятельность внутренних органов, желез внешней и внутренней секреции, кровеносных и лимфатических сосудов. Первые сведения о структуре и функции вегетативной нервной системы принадлежат Галену (II век н. э.). J. Reil (1807) ввёл понятие «вегетативная нервная система», а J. Langley (1889) дал морфологическое описание вегетативной нервной системы, предложил деление её на симпатический и парасимпатический отделы, ввёл термин «автономная нервная система», учитывая способность последней самостоятельно осуществлять процессы регуляции деятельности внутренних органов. В настоящее время в русско, немецко-, франкоязычной литературе можно встретить термин вегетативная нервная система, а в англоязычной– автономная нервная система (АНС). Деятельность вегетативной нервной системы в основном непроизвольна и сознанием непосредственно не контролируется, направлена на поддержание постоянства внутренней среды и приспособление её к изменяющимся условиям внешней среды.
Анатомия вегетативной нервной системы
С точки зрения иерархии управления, вегетативная нервная система условно делят на 4 этажа (уровня). Первый этаж – интрамуральные сплетения, второй – паравертебральные и превертебральные ганглии, третий – центральные структуры симпатической нервной системы (СНС) и парасимпатической нервной системы (ПСНС). Последние представлены скоплениями преганглионарных нейронов в стволе мозга и спинном мозге. Четвёртый этаж включает высшие вегетативные центры (лимбико-ретикулярный комплекс – гиппокамп, грушевидная извилина, миндалевидный комплекс, перегородка, передние ядра таламуса, гипоталамус, ретикулярная формация, мозжечок, кора больших полушарий). Первые три этажа формируют сегментарный, а четвёртый – надсегментарный отделы вегетативной нервной системы.
Кора головного мозга является высшим регуляторным центром интегративной деятельности, активируя как моторные, так и вегетативные центры. Лимбико-ретикулярный комплекс и мозжечок отвечают за координацию вегетативных, поведенческих, эмоциональных, нейроэндокринных реакций организма. В продолговатом мозге расположен сердечно — сосудистый центр, объединяющий парасимпатический (кардиоингибиторный), симпатический (вазодепрессорный) и сосудодвигательный центры, регуляция которых осуществляется подкорковыми узлами и корой головного мозга. Ствол мозга постоянно поддерживает вегетативный тонус. Симпатический отдел вегетативной нервной системы вызывает мобилизацию деятельности жизненно важных органов, повышает энергообразование в организме, стимулирует работу сердца (повышается ЧСС, возрастает скорость проведения по специализированным проводящим тканям, увеличивается сократимость миокарда). Парасимпатический отдел вегетативной нервной системы оказывает трофотропное действие, способствуя восстановлению нарушенного во время активности организма гомеостаза, действует угнетающе на сердце (снижает ЧСС, атриовентрикулярную проводимость и сократимость миокарда).
Ритм сердца определяется способностью специализированных клеток сердца спонтанно активироваться, так называемым свойством сердечного автоматизма. Автоматизм обеспечивает возникновение электрических импульсов в миокарде без участия нервной стимуляции. В нормальных условиях процессы спонтанной диастолической деполяризации, определяющие свойство автоматизма, наиболее быстро протекают в синоатриальном узле (СУ). Именно синоатриальный узел задаёт ритм сердца, являясь водителем ритма 1 порядка. Обычная частота синусового импульсообразования – 60 – 100 импульсов в минуту, т.е. автоматизм синоатриального узла не является постоянной величиной, он может изменяться в связи с возможным смещением водителя ритма в пределах узла. В настоящее время ритм сердца рассматривается не только как показатель собственной функции ритмовождения синоатриального узла, но в большей степени как интегральный маркёр состояния множества систем, обеспечивающих гомеостаз организма. В норме основное модулирующее влияние на ритм сердца оказывает вегетативная нервная система.
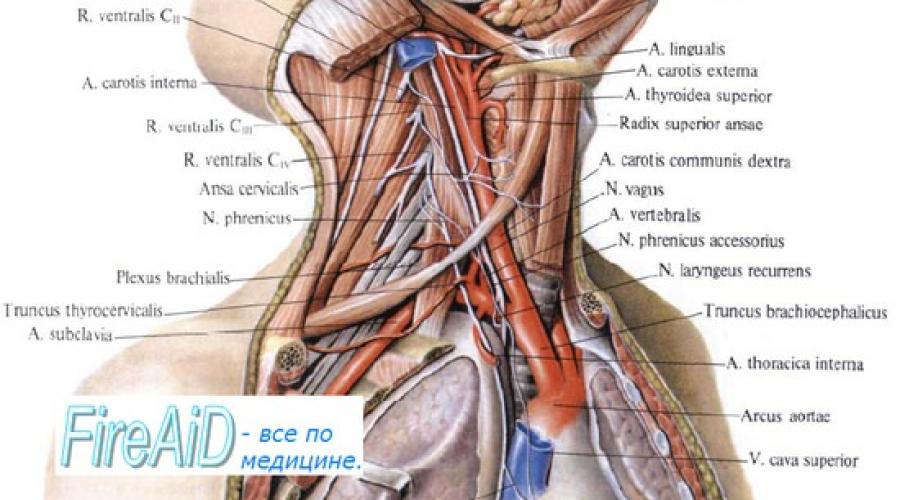
Иннервация сердца
Преганглионарные парасимпатические нервные волокна берут начало в продолговатом мозге, в клетках, которые находятся в дорсальном ядре блуждающего нерва (nucleus dorsalis n. vagi) или двойном ядре (nucleusambigeus) Х черепного нерва. Эфферентные волокна проходят вниз по шее, вблизи общих сонных артерий и через средостение, образуя синапсы с постганглионарными клетками. Синапсы формируют парасимпатические ганглии, располагающиеся внутристеночно, преимущественно вблизи синоатриальных узлов и атриовентрикулярного соединения (АВС). Нейромедиатором, выделяющимся из постганглионарных парасимпатических волокон, является ацетилхолин. При этом раздражение блуждающего нерва приводит к замедлению диастолической деполяризации клеток, снижает частоту сердечных сокращений(ЧСС). При непрерывной стимуляции блуждающего нерва латентный период реакции составляет 50-200 мс, что обусловлено действием ацетилхолина на специфические ацетилхолинергические К+ каналы в клетках сердца.
Постоянный уровень ЧСС достигается через несколько сердечных циклов. Однократная стимуляция блуждающего нерва или короткая серия импульсов оказывает влияние на ЧСС в течение последующих 15-20 с, с быстрым возвращением к контрольному уровню, благодаря быстрой деградации ацетилхолина в области синоатриального узла и атриовентрикулярного соединения. Сочетание 2 характерных особенностей парасимпатической регуляции – короткого латентного периода и быстрого угасания ответной реакции, позволяет ей осуществлять быструю регуляцию и контроль за работой синоатриального узла и атриовентрикулярного соединения практически при каждом сокращении.
Волокна правого блуждающего нерва иннервируют преимущественно правое предсердие и особенно обильно СУ, а левого блуждающего нерва – атриовентрикулярного соединения. В результате при раздражении правого блуждающего нерва более выражен отрицательный хронотропный эффект, а при стимуляции левого – отрицательный дромотропный.
Парасимпатическая иннервация желудочков выражена слабо, в основном представлена в задненижней стенкелевого желудочка. Поэтому при ишемии или инфаркте миокарда данной области отмечается брадикардия и гипотония, обусловленные возбуждением блуждающего нерва и описаны в литературе как рефлекс Бецольда Яриша.
Преганглионарные симпатические волокна берут начало в интермедиалатеральных столбах 5-6 верхних грудных и 1-2 нижних шейных сегментах спинного мозга. Аксоны преганглионарных и постганглионарных нейроновобразуют синапсы в трёх шейных и звёздчатом ганглиях.
В средостении постганглионарные волокна симпатических и преганглионарные волокна парасимпатических нервов соединяются вместе, образуя сложное нервное сплетение смешанных эфферентных нервов, идущих к сердцу. Постганглионарные симпатические волокна достигают основания сердца в составе адвентиции крупных сосудов, где образуют обширное сплетение эпикарда. Затем они проходят сквозь миокард, вдоль коронарных сосудов. Нейромедиатором, выделяющимся из постганглионарных симпатических волокон, является норадреналин, уровень которого одинаков как в СУ, так и в области правого предсердия.
Повышение симпатической активности вызывает увеличение ЧСС, ускоряет диастолическую деполяризацию клеточных мембран, смещает водитель ритма к клеткам с самой высокой автоматической активностью. Пристимуляции симпатических нервов ЧСС повышается медленно, латентный период реакции составляет 1-3 с, а установившийся уровень ЧСС достигается лишь через 30-60 с от начала стимуляции. На скорость реакции влияет то, что медиатор вырабатывается довольно медленно нервными окончаниями, а воздействие на сердце осуществляется через относительно медленную систему вторичных мессинджеров – аденилатциклазу. После прекращения стимуляции хронотропный эффект исчезает постепенно. Скорость исчезновения эффекта стимуляции определяется снижением концентрации норадреналинав межклеточном пространстве, которая изменяется путём поглощения последнего нервными окончаниями, кардиомиоцитами и диффузией нейромедиатора в коронарный кровоток. Симпатические нервы практически равномерно распределены по всем отделам сердца, с максимальной иннервацией области правого предсердия. Симпатические нервы правой стороны преимущественно иннервируют переднюю поверхность желудочков и СУ, а левой – заднюю поверхность желудочков и атриовентрикулярного соединения.
Афферентная иннервация сердца осуществляется в основном миелинизированными волокнами, идущими в составе блуждающего нерва. Рецепторный аппарат в основном представлен механо- и барорецепторами, расположенными в правом предсердии, в устьях легочных и полых вен предсердий, желудочках, дуге аорты, синокаротидном синусе. По мнению большинства исследователей, регуляторные влияния ПСНС на СУ и атриовентрикулярного соединения значительно превосходят влияния СНС.
Деятельность ВНС находится под влиянием центральной нервной системы (ЦНС) по механизму обратной связи. Обе системы тесно связаны между собой, а нервные центры на уровне ствола и полушарий головного мозга невозможно разделить морфологически. Самый верхний уровень взаимодействия осуществляется в сосудодвигательном центре, куда поступают и где обрабатываются афферентные сигналы из сердечно — сосудистой системы и где происходит регуляция эфферентной активности симпатической и парасимпатической нервной деятельности. Кроме интеграции на уровне ЦНС, важную роль играет также взаимодействие на уровне пре- и постсинаптических нервных окончаний, что подтверждено результатами анатомических и гистологических исследований. В последних исследованиях обнаружены особые клетки, содержащие большие запасы катехоламинов, на которых расположены синапсы, образованные терминальными окончаниями блуждающего нерва, что указывает на возможность прямого воздействия блуждающего нерва на адренергические рецепторы. Установлено, что часть внутри сердечных нейроцитов имеют положительную реакцию на моноаминоксидазу, что указывает на их роль в метаболизме норадреналина.
Несмотря на разнонаправленное в целом действие СНС и ПСНС, при одновременной активации обоих отделов ВНС их эффекты не складываются простым алгебраическим способом и взаимодействие нельзя выразить линейной зависимостью. В литературе описано несколько типов взаимодействия отделов ВНС. Согласно принципу «акцентированного антагонизма», ингибирующий эффект данного уровня парасимпатической активности тем сильнее, чем выше уровень симпатической активности, и наоборот. С другой стороны, при достижении определённого результата снижения активности в одном отделе ВНС происходит повышение активности другого отдела по принципу «функциональной синергии». При исследовании вегетативной реактивности необходимо учитывать «закон исходного уровня»,согласно которому чем выше исходный уровень, тем в более деятельном и напряженном состоянии находится система, тем меньший ответ возможен при действии возмущающих стимулов.
Состояние отделов ВНС претерпевает значительные изменения на протяжении жизни человека. В младенческом возрасте отмечается значительное преобладание симпатических нервных влияний при функциональной иморфологической незрелости обоих звеньев ВНС. Развитие симпатического и парасимпатического отделов ВНС после рождения происходит интенсивно, и к моменту полового созревания плотность расположения нервных сплетений в различных отделах сердца достигает наивысших показателей. При этом у лиц молодого возраста отмечается доминирование парасимпатических влияний, проявляющихся в исходной ваготонии в состоянии покоя.
Начиная с 4-го десятилетия жизни, начинаются инволютивные изменения в аппарате симпатической иннервации, при сохранении плотности холинергических нервных сплетений. Процессы десимпатизации приводят к снижению симпатической активности и падению плотности распределения нервных сплетений на кардиомиоцитах, гладкомышечных клетках, способствуя гетерогенности потенциал зависимых свойств мембраны в клетках проводящей системы, рабочем миокарде, стенках сосудов, гиперчувствительности рецепторного аппарата к катехоламинам и могут служить основой аритмий , в том числе и фатальных. Имеются также и половые различия в состоянии вегетативного нервного тонуса.
Так, у женщин молодого и среднего возраста (до 55 лет) отмечена более низкая активность симпатической нервной системы, чем у мужчин аналогичного возраста. Таким образом, вегетативная иннервация различных отделов сердца неоднородна и несимметрична, имеет возрастные и половые различия. Согласованная работа сердца является результатом динамического взаимодействия отделов ВНС между собой.
Рефлекторная регуляция сердечной деятельности
Артериальный барорецепторный рефлекс является ключевым механизмом краткосрочной регуляции артериального давления (АД). Оптимальный уровень системного артериального давления является одним из наиболее важных факторов, необходимых для адекватной работы сердечно — сосудистой системы. Афферентные импульсы от барорецепторов каротидных синусов и дуги аорты по ветвям языкоглоточного нерва (IX пара) и блуждающего нерва (Х пара) поступают к кардиоингибиторному и сосудодвигательному центру продолговатого мозга и другим отделам ЦНС. Эфферентное плечо барорецепторного рефлекса образуется симпатическими и парасимпатическими нервами. Импульсация от барорецепторов повышается на увеличении абсолютной величины растяжения и скорости изменения растяжения рецепторов.
Повышение частоты импульсации от барорецепторов оказывает тормозящее влияние на симпатические центры и возбуждающее на парасимпатические, что приводит к снижению вазомоторного тонуса в резистивных и емкостных сосудах, уменьшению частоты и силы сердечных сокращений. Если среднее АД резко снижается, тонус блуждающего нерва практически исчезает, арефлекторная регуляция осуществляется исключительно за счёт изменений эфферентной симпатической активности. При этом повышается общее периферическое сопротивление сосудов, увеличивается частота и сила сердечных сокращений, направленных на восстановление исходного уровня АД. И наоборот, если АД резко повышается, симпатический тонус полностью угнетается, а градация рефлекторной регуляции происходит только благодаря изменениям эфферентной регуляции вагуса.
Повышение давления в желудочках вызывает раздражение субэндокардиальных рецепторов растяжения и активацию парасимпатического кардиоингибиторного центра, что приводит к рефлекторной брадикардии и вазодилатации. Рефлекс Бейбриджа характеризуется повышением симпатического тонуса с повышением ЧСС в ответ на увеличение внутрисосудистого объема крови и повышение давления в крупных венах и правом предсердии.
При этом происходит рост ЧСС, несмотря на сопутствующий подъём АД. В реальной жизни рефлекс Бейбриджа преобладает над артериальным барорецепторным рефлексом в случае увеличения объёма циркулирующей крови. Исходно и при уменьшении объёма циркулирующей крови барорецепторный рефлекс преобладает над рефлексом Бейбриджа.
Ряд факторов, участвующих в поддержании гомеостаза организма, влияет на рефлекторную регуляцию сердечной деятельности, при отсутствии значимых изменений активности ВНС. К ним относятся хеморецепторный рефлекс, изменения уровня электролитов крови (калия, кальция). На частоту сердечных сокращений оказывают влияние также фазы дыхания: вдох вызывает угнетение блуждающего нерва и ускорение ритма, выдох – раздражение блуждающего нерва и замедление сердечной деятельности.
Таким образом, в обеспечении вегетативного гомеостаза участвует большое количество разнообразных регуляторных механизмов. По мнению большинства исследователей, ритм сердца является не только показателем функции СУ, но и интегральным маркёром состояния множества систем, обеспечивающих гомеостаз организма, с основным модулирующим влиянием ВНС. Попытка выделить и количественно оценить влияние на ритм сердца каждого из звеньев – центрального, вегетативного, гуморального, рефлекторного – несомненно, является актуальной задачей в кардиологической практике, так как её решение позволит разработать дифференциально-диагностические критерии сердечно — сосудистой патологии на основании простой и доступной оценки состояния ритма сердца.
Б. Лаун и Р. Л. ВерьеРЕФЕРАТ. Увеличение тонуса парасимпатической нервной системы, вызванное либо стимуляцией вагуса, либо прямым воздействием на мускариновые рецепторы, значительно уменьшает склонность миокарда нормальных и ишемизированных желудочков к развитию фибрилляций. Этот защитный эффект является результатом антагонистического взаимодействия реакций миокарда на повышение нервной и гуморальной активности, влияющих на порог возникновения фибрилляций желудочков: Эти механизмы функционируют как у бодрствующего, так и у анестезированного животного. Полученные результаты, несомненно, имеют большое значение для клинической практики.
ВВЕДЕНИЕ
Вопрос о влиянии парасимпатической нервной системы на возбудимость клеток миокарда желудочков постоянно подвергается переоценке. В настоящее время общепринято, что вагусная иннервация не распространяется на миокард желудочков. С точки зрения клинициста, очевидно, что хотя холинергическое воздействие может оказать влияние на тахикардию, тем не менее место приложения ацетилхолина расположено вне желудочков. С другой стороны, проведенные в последнее время исследования позволяют утверждать, что воздействие со стороны парасимпатической нервной системы может изменять электрические свойства миокарда желудочков . Как было показано несколькими группами исследователей, стимуляция вагуса существенно влияет на возбудимость клеток желудочка и их склонность к фибрилляции . Эти эффекты могут быть опосредованы наличием богатой холинергической иннервации специализированной проводящей системы сердца, которая была обнаружена как в сердце собаки, так и в сердце человека .
Нами было показано, что влияние вагуса на вероятность возникновения фибрилляций желудочков (ФЖ) зависит от фонового уровня тонуса симпатических нервов сердца . Это положение вытекает из ряда экспериментальных наблюдений. Например, влияние вагуса возрастает у торакотомированных животных, у которых проявляется повышенный симпатический тонус, а также во время стимуляции симпатических нервов и инъекции катехоламинов. Такое действие вагуса на склонность желудочков к фибрилляции устраняется при блокаде |3-ре^ цепторов.
До сих пор точно не установлено, способна ли парасимпатическая нервная система изменять склонность желудочков к фибрилляции, развивающейся во время острой ишемии миокарда. Kent и Epstein с.соавт показали, что стимуляция вагуса значительно увеличивает порог ФЖ и уменьшает склонность ишемизированного сердца собаки к фибрилляции. Согг в. Gillis с соавт. обнаружили, что наличие интактных вагусных нервов предупреждает развитие ФЖ во время перевязки левой передней нисходящей артерии сердца наркотизированной хлоралозой кошки, но не дает никаких преимуществ при перевязке правой коронарной артерии. Yoon с соавт. и James с соавт. не смогли выявить какое-либо влияние стимуляции вагуса на порог ФЖ во время окклюзии левой передней нисходящей коронарной артерии собаки. Согг с соавт. даже обнаружили, что стимуляция парасимпатической нервной системы скорее усиливает, чем ослабляет, аритмии, которые возникают при снятии лигатуры с артерии, сопровождаемой реперфузией ишемизироваиного миокарда.
К этому также относится нерешенная проблема, модулирует ли тоническая активность парасимпатической нервной системы электрическую устойчивость клеток желудо"чка животного, находящегося в ненаркотизированном состоянии. Данные, полученные на наркотизированных животных при стимуляции нервов или введения лекарств, представляют собой ценную информацию, однако такие подходы в какой-то мере артефактны, и результаты требуют подтверждения на ненаркотизированном интактном организме. До последнего времени исследования животных в бодрствующем состоянии с такой целью не проводились в связи с отсутствием подходящих биологических моделий для оценки склонности миокарда к ФЖ. Однако эта трудность была преодолена, когда в "качестве надежного показателя склонности сердца к ФЖ использовали порог повторных экстравозбуждений, что позволило в результате отказаться от необходимости вызывать ФЖ и проводить сопутствующие реанимационные процедуры .
Задачи настоящего исследования заключались в следующем: 1) изучить влияние стимуляции вагуса и прямой активации метахолииом мускариновых рецепторов на склонность сердца к ФЖ во время острой ишемии миокарда и при репер-фузии, 2) определить, изменяет ли тоническая активность парасимпатической нервной системы склонность желудочков к фибрилляции при ненаркотизированном состоянии животного, и 3) оценить, имеют ли полученные на животных данные-какое-либо отношение к клиническим задачам.
МАТЕРИАЛ И МЕТОДЫ
Исследования на наркотизированных животных
Общие процедуры
Исследования были выполнены на 54 здоровых беспородных собаках массой от 9 до 25 кг. Не менее чем за 5 дней до-исследования под общим пентобарбитуратным наркозом проводили вскрытие грудной клетки с левой стороны в четвертом" межреберном пространстве. После обнажения сердца вокруг левой передней нисходящей артерии на уровне ушка левого" предсердия помещали баллончик, связанный с катетером и предназначенный для окклюзии. Катетер выводили под кожей наружу на затылке.
В день исследования собак наркотизировали с помощью а-хлорало"зы 100 мг/кг внутривенно. Искусственное дыхание поддерживали через эндотрахеальную трубку, соединенную с насосом Harvard, подающим смесь комнатного воздуха со 100% кислородом.Подачу кислорода в смеси осуществляли таким образом, чтобы артериальное рО2, находилось между 125 и 225 мм рт. ст. рН артериальной крови поддерживали в диапазоне от 7,30 до 7,55. Артериальное давление в брюшной аорте изменяли с помощью катетера, введенного через бедренную артерию и присоединенного к датчику давления Statham P23Db. Электрограмму (ЭГ) правого желудочка регистрировали с помощью монополярного внутриполостного отведения.
Исследование сердца
В течение всего эксперимента при помощи стимуляции правого желудочка поддерживали постоянный ритм сердца. Для поддержания искусственного ритма и нанесения тестирующих стимулов использовали биполярный катетер (Medtronic №5819), введенный через правую яремную вену и помещенный под флюороскопическим контролем в районе верхушки правого желудочка. Поддержание искусственного ритма достигалось "стимулами, амплитуда которых на 50-100% была выше порога, межстимуляциотаный интервал составлял от 333 до 300 мс, что соответствует частотам возбуждения желудочка от 180 до 200 в минуту.
Порог фибрилляции желудочков определяли с помощью одиночного стимула длительностью 10 мс. Это определение состояло в следующем: электрическую диастолу исследовали с помощью импульса 4 мА с интервалом 10 мс, начиная от конца эффективного рефрактерного периода до завершения Г-волны. Затем величину тока увеличивали с шагом 2 мА и при такой величине стимула продолжали исследование диастолы в течение 3 с. Наименьшую интенсивность стимула, вызывающую ФЖ, принимали в качестве порога ФЖ.
Использовали следующий протокол эксперимента: полная окклюзия левой передней нисходящей коронарной артерии достигалась надуванием заранее имплантированного катетера с баллоном и продолжалась в течение 10 мин. Во время окклюзии порог ФЖ оценивали с минутным интервалом. Через 10 мин после начала окклюзии резко уменьшали давление в баллоне и снова определяли порог ФЖ. Осуществляли две окклюзии с экспериментальным исследованием и без него, разделенных интервалом по крайней мере 20 мин .
Дефибрилляцию производили обычно за 3 с с помощью импульса постоянного тока, получаемого при разряде конденштора с энергоемкостью 50-100 Вт"С от дефибриллятора.11 лупа. Эта реанимационная процедура существенно не влияет на стабильность порога ФЖ.
Стимуляция вагуса
Шейный вагосимпатический ствол перерезали с двух сторон на 2 см ниже места бифуркации сонной артерии. К дисталь-иым концам перерезанного нерва прикрепляли изолированные биполярные электроды. Раздражение нерва производили с помощью прямоугольных импульсов длительностью 5 мс и напряжением 3-15 В при частоте стимуляции 20 Гц. Амплитуду раздражающих импульсов подбирали таким образом, чтобы при независимом раздражении либо правого, либо левого ство-дов вагуса достигалась остановка сердца. Порог фибрилляции желудочков определяли до, во время и после двусторонней стимуляции вагуса. Частоту сердечного ритма во время определения порога ФЖ постоянно искусственно поддерживали на уровне 200 ударов в минуту.
Введение метахолина
Внутривенное введение мускаринового агониста - хлорида ацетил-(Б,Ь)-бета-метилхолина (J. Т. Baker Company) в физиологическом растворе осуществляли со скоростью 5 мкг/ (кг-мин), используя инфузионный насос «Harvard». Максимальный эффект на порог ФЖ достигался через 30 мин после начала введения; в этот момент начинали проведение всей последовательности тестирований с окклюзией коронарной артерии и реперфузией. Введение вещества продолжалось в течение всего исследования.
ИССЛЕДОВАНИЯ НА БОДРСТВУЮЩИХ ЖИВОТНЫХ
Исследования проводили на 18 взрослых беспородных собаках массой от 10 до 15 кг.
Для проведения обратимой холодовой блокады парасимпатической активности нервов сердца был разработан специальный метод. Для этого выделяли часть вагосимпатического ствола длиной 3-4 см и помещали его на шее в кожную трубку. Таким образом, по обе стороны шеи были созданы «вагусные петли», которые отделяли изолированные сегменты нервов от других шейных структур. Это позволяло поместить вокруг вагусных петель охлаждающие наконечники для того, чтобы произвести обратимую блокаду нервной активности.
Относительный вклад активности вагусных афферентов и эфферентов в эффект, производимый охлаждением, определяли путем сравнения результатов, полученных при охлаждении вагуса с селективной блокадой вагусных эфферентов при внут-ривенно-м введении атропина.
Исследование сердца:
Для изучения склонности сердца к ФЖ использовали метод определения порога повторных экстравозбуждений (ПЭ) как описано ранее . Вкратце, порог склонности к ФЖ оценивался следующим образом: при поддержании постоянной частоты сердечного ритма 220 ударов в минуту сканирование повторным стимулом для определения порога ПЭ осуществляли при интенсивности стимула, равной двойному значению порога в середине диастолы, начиная с 30 мс после окончания рефрактерного периода. Тестирующий стимул подавали с каждым разом все раньше с шагом 5 мс, пока не подходили к концу рефрактерного периода. Если при этом не возникали ПЭ, амплитуду стимула увеличивали на 2 мА и повторяли процесс сканирования. Порог ПЭ считали равным минимальному значению тока, при котором ПЭ возникали в двух из каждых трех попыток. Порог ПЭ принимали в качестве порога уязвимости OK ФЖ.
Психологические условия
Для изучения влияния симпатических - парасимпатических взаимодействий в состоянии бодрствования собак помещали в стрессогенные условия, которые увеличивают поступление в сердце адренергических агониетов .
Стрессогенные условия заключались в закреплении собаки в станке Павлова, что вызывало ограничение двигательных возможностей. К сердечным катетерам подключали кабели для непрерывного наблюдения за ЭГ, подачи стимулов от искусственного водителя ритма и тестирующих стимулов. Отдельный удар электрическим током длительностью 5 мс осуществляли от дефибриллятора через медные пластины (80 см2), прикрепленные к грудной клетке. Собак оставляли в ремнях на 10 мин до нанесения электрического удара и еще на 10 мин после подачи тока. Процедуру повторяли 3 дня подряд. На 4-й день нанесения электрического удара исследовали влияние стрессогенных условий содержания на пороговый период уязвимости сердца к ФЖ до и во время блокады вагусных эфферентов атропином (0,05 мг/кг).
РЕЗУЛЬТАТЫ
15л и ниие стимуляции холинергических нервов на склонность сердца к ФЖ во время ишемии 1миокарда и при реперфузии
Изучение влияния стимуляции вагуса на порог ФЖ до и и<> время 10-минутного периода окклюзии передней левой нисходящей коронарной артерии с последующим внезапным иоостановлением кровотока было проведено на 24 собаках, наркотизированных хлоралозой. В отсутствие стимуляции вагуса окклюзия коронарной артерии и реперфузия приводили к значительному снижению порога фибрилляции (рис. 1), Снижение порога происходило в первые 2 мин после окклюзии и продолжалось от 5 до 7 мин. Затем порог быстро возвращался к значению, наблюдаемому в контроле до окклюзии. После восстановления проводимости коронарной артерии падение порога происходило почти мгновенно - за 20-30 с, но продолжалось недолго - менее 1 мин. Стимуляция вагуса значительно повышала порог ФЖ до окклюзии коронарной артерии (от 17±2 мА до З3.±4 мА, р<0,05) и уменьшала снижение порога, связанное с ишемией миокарда (18±4 мА по сравнению с 6±1 мА без стимуляции, р<С0,05). Во время реперфузии никакого защитного действия стимуляции вагуса не обнаружено (3±1 мА по сравнению с 5±1 мА без стимуляции).
Влияние селективной "Стимуляции мускариновых рецепторов с помощью метахолина на уязвимость сердца к ФЖ исследовали на 10 собаках. Введение метахолина приводило к результатам, качественно аналогичным тем, которые были получены при стимуляции вагуса. Так, метахолин повышал порог ФЖ до и во время окклюзии коронарной артерии, но был неэффективен при падении порога, связанном с реперфузи-ivii (рис. 2).
Влияние активности вагуса на склонность сердца
и спонтанным ФЖ при ишемии миокарда и реперфузии
Исследование влияния стимуляции вагуса на появление спонтанной ФЖ при окклюзии левой передней нисходящей коронарной артерии и артерии межжелудочковой перегородки было проведено дополнительно на 16 собаках. С помощью искусственной стимуляции желудочка поддерживали постоянную частоту сердечного ритма, равную 180 уд/мин. В отсутствие стимуляции вагуса окклюзия коронарной артерии вы-нвала ФЖ у 7 из 10 собак (70%), в то время как при одновременной стимуляции вагуса спонтанная ФЖ при окклюзии
Этот вопрос был изучен на 10 бодрствующих собаках, у которых оба вагуса были хронически выделены на шее в кожные трубки. Импульсацию в вагосимпатическом стволе обратимо блокировали при помощи охлаждающих наконечников, помещенных вокруг кожных вагусных петель. Холодовая блокада левой и правой вагусных петель увеличивала частоту сердечного ритма с 95+5 ударов в минуту до 115±7 и 172+ + 16 ударов в минуту соответственно. Когда обе вагусные петли были охлаждены одновременно, частота сердечного ритма увеличилась до 208+20 ударов в минуту. Все изменения частоты сердечного ритма были статистически достоверны с р< 0,01 (рис. 4).
Исследование влияния селективной блокады вагусных эф-! ферентов с помощью атропина на порог ПЭ было проведено на 8 бодрствующих собаках, содержавшихся в стрессогенных условиях, создаваемых с помощью иммобилизации в станке Павлова с нанесением чрезкожного удара электрическим током средней тяжести. До выключения воздействия на сердце вагусной импульсации порог ПЭ составлял 15+1 мА. При введении атропина (0,05 мг/кг) порог значительно снизился и составил 8±1 мА (снижение на 47%, р<0,0001) (рис. 5).
Этот эффект развивался независимо от изменений сердечного ритма, так как частота сердечного ритма поддерживалась постоянной на уровне 200 ударов в минуту в течение всего времени проведения электрического тестирования. Блокада вагуса с помощью атропина несущественно влияла на порог ПЭ у собак, содержавшихся в клетках с нестреосогенными условиями (22+2 мА и 19+3 мА до и при действии вещества соответственно).
ОБСУЖДЕНИЕ
В настоящее время накоплено значительное количество данных, указывающих на наличие прямого влияния парасимпатической нервной системы на хронотропные и изотропные свойства и возбудимость миокарда желудочков. Значительно меньше доказано, является ли величина этого влияния достаточной, чтобы объяснить некоторое защитное действие от возникновения ФЖ активности холинергических нервов в ише-мизированном сердце. Кроме того, мало известно о значении активности парасимпатических нервов в склонности сердца к ФЖ в двух различных условиях, которые, возможно, играют важную роль в возникновении внезапной смерти у человека а именно при внезапной окклюзии коронарной артерии и восстановлении ее проходимости с реперфузией ишемизиро-ванной области. До сих пор не определено значение тонической активности вагуса для уменьшения склонности к ФЖ. Еще один нерешенный вопрос состоит в том, может ли такая тоническая активность парасимпатической нервной системы влиять на склонность желудочков к фибрилляции при слабых психофизиологических стрессах. Настоящее исследование проливает некоторый свет на эти вопросы.
Эффект стимуляции вагуса во время ишемии миокарда и при реперфузии
Мы установили, что интенсивная парасимпатическая активность, возникающая при электрическом раздражении децентрализованного вагуса, или прямая стимуляция мускарино-вых рецепторов с помощью метахолина уменьшает склонность сердца собаки к ФЖ во время острой ишемии миокарда. Это также подтверждается наблюдениями, показывающими, что увеличение холинертичеокой активности значительно уменьшает падение порога ФЖ и склонность к спонтанным ФЖ во время окклюзии коронарной артерии. Эти эффекты не связаны с изменением сердечного ритма, так как его частоту поддерживали на постоянном уровне с помощью искусственного водителя ритма. Ни стимуляция вагуса, ни активация мускари-новых рецепторов не оказывали никакого положительного действия во время реперфузии.
Что же обусловливает различное влияние парасимпатической нервной системы на порог ФЖ во время ишемии миокарда и во время реперфузии? Предполагают, что склонность сердца к ФЖ при окклюзии "коронарной артерии и при реперфузии обусловлена различными механизмами . Вероятно, основную роль в увеличении склонности сердца к ФЖ во время острой окклюзии коронарной артерии играет рефлекторная активация симпатической нервной системы в сердце . Эту гипотезу подтверждает то, что изменение в поступлении адренергических веществ в сердце хорошо коррелирует с развитием во времени снижения порога ФЖ и появлением спонтанных ФЖ при окклюзии коронарной артерии . Если воздействие симпатических аминов на миокард уменьшено хирургическими или фармакологически-iin методами , то при этом достигается значительный защитный эффект против вызванных ишемией ФЖ. Таким образом, активность парасимпатической нервной системы уменьшает склонность сердца к ФЖ во время окклюзии коронарной артерии "благодаря противодействию профибриллятор-ному влиянию увеличенной адренергилеской активности. Такой положительный эффект увеличения холинергической активности может быть следствием ингибирования освобождения норадреналипа из симпатических нервных окончаний либо следствием уменьшения реакции рецепторов на воздействие катехоламинов .
Однако увеличение склонности миокарда к фибрилляции во время реперфузии, по-видимому, обусловлено неадренер-гическими факторами. Имеющиеся в настоящее время данные указывают на то, что это явление может быть связано продуктами метаболизма, вымываемыми в кровь при клеточной ишемии и некрозе . Было показано, что если кровоток в ишемическом миокарде восстанавливается постепенно или если перфузия производится раствором, лишенным кислорода, частота случаев появления желудочковых аритмий при восстановлении кровотока существенно снижается . Наблюдения, показывающие, что ФЖ возникает в течение нескольких секунд после внезапного восстановления коронарного артериального кровотока, также указывают на участие в этом процессе вымываемых из поврежденной зоны продуктов метаболизма . Предотвращение воздействия симпатических веществ на сердце с помощью хирургического или фармакологического вмешательства оказывается неэффективным для предупреждения ФЖ при восстановлении кровотока. А так как холинергические агонисты проявляют свое защитное влияние лишь через антиадренергическое действие, это может частично объяснить их неспособность уменьшить склонность миокарда к ФЖ во время реперфузии.
Сильное влияние активности парасимпатической нервной системы на частоту сердечного ритма может существенно изменить действие стимуляции вагуса на склонность желудочка к аритмиям. Например, Kerzner с соавт. показали, что стимуляция вагуса не полностью подавляет аритмии, возникающие при инфаркте миокарда. Напротив, эти исследователи обнаружили, что увеличение активности парасимпатической нервной системы или введение ацетилхолина неизменно вызывает желудочковую тахикардию во время спокойной без аритмий фазы инфаркта миокарда у собак. Такое аритмоген-ное действие полностью зависит от частоты сердечного ритма и может быть предупреждено с помощью искусственного водителя ритма.
Влияние тонической активности парасимпатической нервной системы на склонность желудочков к фибрилляции у животных, находящихся в бодрствующем состоянии
Результаты настоящего исследования указывают на то, ч:то в покое в состоянии бодрствования собаки ее сердце испытывает значительное тоническое влияние парасимпатической нервной системы. Холодовая блокада либо правого, либо левого вагуса приводит к существенным изменениям частоты сердечного ритма; однако эффект более выражен при блокаде правого вагуса (см. рис. 4). Это соответствует тому, что правый вагус оказывает преобладающее воздействие на синоат-риальный узел с некоторым наложением влияния от левого «агуса . Таким образом, максимальное увеличение частоты сердечного ритма возникает при одновременном охлаждении правого и левого вагусных нервов.
Установив, что тоническая активность парасимпатической нервной системы оказывает значительное влияние на пей-смекерную ткань, имеет смысл исследовать, можно ли выявить какое-либо влияние активности вагуса на электрические свойства желудочка. В этих экспериментах для селективной блокады активности вагусных эфферентов использовали атропин. Собак помещали в станок Павлова для иммобилизации с целью повышения симпатического влияния на сердце . Такая планировка эксперимента позволяла изучать влияние взаимодействия симпатических и парасимпатических реакций на склонность миокарда к ФЖ у бодрствующих животных. Нами установлено, что введение относительно низких доз атропина (0,05 мг/кг) приводит почти к 50% снижению порога фибрилляции желудочков. Это позволяет сделать вывод, что значительная тоническая активность вагуса у бодрствующего животного, содержащегося в стрессогенных условиях, ча-стично ослабляет профибрилляторное влияние эверсивных психофизиологических стимулов.
Кроме того, при использовании такой экспериментальной схемы защитное действие вагуса скорее всего обусловлено антагонистическим к адренергическому механизму действием. Это предположение подтверждается двумя типами наблюдений. Во-первых, наши предыдущие исследования показали, что склонность миокарда к фибрилляции в такой модели стрессогенных услоиий тесно коррелирует с уровнем циркулирующих в крови катехоламинов и что предупреждение симпатического влияния на сердце либо с помощью бета-блокады, либо при симпатэктомии существенно снижает вызванное стрессогенными условиями увеличение склонности к фибрилляции . Во-вторых, наблюдения De Silva с соавт. показывают, что увеличение тонического воздействия парасимпатической нервной системы при введении морфина собакам, находящимся в стрессогенных условиях иммобилизации, по-... вышает порог ФЖ до величины, наблюдаемой при отсутствии стрессорных воздействий. Когда активность вагусных эфферентов блокируется атропином, основная часть защитного действия морфина исчезает. Введение морфина в нестрессо-генных условиях не способно изменить порог ФЖ, видимо, потому, что в этих условиях адренергичеокое влияние на сердце слабое.
Эти данные указывают, что активация блуждающих нервов независимо от того, возникает ли она спонтанно или вызвана фармакологическим агентом, имеет защитное действие на миокард, снижая его склонность к ФЖ при стрессе. Это благотворное влияние скорее всего обусловлено антагонистическим влиянием повышенной активности парасимпатической нервной системы на эффект увеличения адренергической активности в сердце.
КЛИНИЧЕСКОЕ ПРИМЕНЕНИЕ
Более 40 лет назад было показано, что введение холинергического вещества - хлорида ацетил-бета-метилхолина, предупреждает желудочковые аритмии, вызванные у человека введением адреналина . В последнее время в ряде исследований сообщалось, что воздействия, аналогичные активации парасимпатической нервной системы, как, например, стимуляция каротидного синуса или введение ваготониче-ских агентов , снижают частоту желудочковых экстрасистол и предупреждают желудочковую тахикардию. Так как сердечные гликозиды увеличивают тоническое влияние блуждающего нерва на сердце, мы использовали это действие дигиталиса для подавления желудочковых аритмий . Однако в этой клинической области требуются дальнейшие исследования.
Это исследование было проведено Научно-исследовательской лабораторией сердечно-сосудистых заболеваний Гарвардской школы здравоохранения, Бостон, штат Массачусетс. Оно было также поддержано субсидией МН-21384 Национального института психического здоровья и субсидией HL-07776 Национального института сердца, легких и крови Национальных институтов здоровья, Бетезда, штат Мэриленд.
СПИСОК ЛИТЕРАТУРЫ
1. Kent К . М ., Smith Е . R., Redwood D. R. et al. Electrical stability of acu-
tely ischemic myocardium: influences of heart rate and vagal stimulation.-Circulation, 1973, 47: 291-298.
2. Kent K. M., Epstein S. E., Cooper T. et al. Cholinergic innervation of the
canine and human ventricula conducting system: anatomic and elec-trophysiologic correlation.-Circulation, 1974, 50: 948-955.
3. Kolman B. S-, Verrier R. L., Lown B. The effect of vagus nerve stimula-
tion upon vulnerability of the canine ventricular. Role of cympathetic-parasympathetic interactions.-Circulation, 1975, 52: 578-585.
4. Weiss Т ., Lattin G. M., Engelman K. Vagally mediated supression of pre-
mature ventricular contractions in man.-Am. Heart J., 1977, 89: 700- 707.
5. Waxman M. В ., Wald R. W. Termination of ventricular tacycardia by an
increase in cardiac vagal drive.-Criculation, 1977, 56: 385-391.
6. Kolman B. S., Verrier R. L., Lown B. Effect of vagus nerve stimulation
upon excitability of the canine ventricle: role of sympathetic-parasympa-thetic interactions.-Am. J. Cardiol., 1976, 37: 1041-1045.
7. loon M. S., Han J., Tse W. W. et al Effects of vagal stimulation, atropine,
and propranolol on fibrillation threshold of normal and ischemic ventricles.-Am. Heart J., 1977, 93: 60-65.
8. Lown В ., Verrier R. L. Neural activity and ventricular fibrillation.-New
Engl. J. Med., 1976, 294: 1165-1170.
9. Coor P. В ., Gillis R. A. Role of the vagus in the cardiovascular chenges
induced by coronary occlusion.- Circulation 1974, 49: 86-87.
10. Coor P. В ., Pearle D. L., Gillis R. A. Coronary occlusion site as a determi
nant of the cardiac rhythm effects of atropine and vagotomy.-Am. He
art J., 1976, 92: 741-749.
11. James R. G. G., Arnold J. M. O., Allen 1. D. et al. The effects of heart
rate, myocardial ischemia and vagal stimulation on the threshold for ventricular fibrillation.-Circulation, 1977, 55: 311-317.
12. Corr P. В ., Penkoske P. A., Sobel В . Е . Adrenergic influences on arrhyrh-
mias due to coronary occlusion and reperfusion.-Br. Heart J., 1978, 40 (suppl.), 62-70.
13. Matta R. J., Verrier R. L., Lown B. The repetitive extrasystole as an in
dex of vulberability to ventricular fibrillation.-Am. J. Physiol., 1976,
230: 1469-1473.
14. Lown В ., Verrier R. L., Corbalan R. Psychologic stress and threshold
for repetitive ventricular response.-Science, 1973, 182: 834-836.
15. Axelrod P. J., Verrier R. L., Lown B. Vulnerability to ventricular fibril-
lation during acute coronary arterial occlusion and release.-Am. J. Car-diol, 1976, 36: 776-782.
16. Corbalan R., Verrier R. L., Lown B. Differing mechanisms for ventricular
vulnerability during coronary artery occlusion and release.-Am. Heart
Т ., 1976, 92: 223-230.
17. DeSilva R. A., Verrier R. L., Lown B. Effect of psycholofic stress and
sedation with morphine sulfate on ventricular vulnerability.-Am. Heart J., 1978, 95: 197-203.
18. Liang В ., Verrier R. L, Lown B. et al. Correlation between circulation
catecholamme levels and ventricular vulnerability during psychologic stress in conscius dogs.-Proc. Soc. Exp. Biol. Med., 1979, 161:266- 269.
19. Malliani A., Schwartz P. L, Zanchetti A. A sympathetic reflex elicited by
experimental coronary occlusion.-Am. J. Physiol., 1969, 217: 703-709.
20. Kelliher G. ]., Widmer C, Roberts J. Influence of the adrenal medulla
on cardiac rhythm disturbances following acute coronary artery occlu
sion.-Recent. Adv. Stud. Cardiac. Struct. Metab.; 1975, 10: 387-400.
21. Harris A. S., Otero H., Bocage A. The induction of arrhythmias by sym
pathetic activity before and after occlusion of a coronary artery in the
canine heart.-J. Electrocardiol., 1971, 4: 34 -43.
22. Khan M. L, Hamilton J. Т ., Manning G. W. Protective effects of beta-
adrenoceptor blockade in experimental occlusion in conscious dogs.- Am. J. Cardiol., 1972, 30: 832-837.
23. Levy M. N., Blattberg B. Effect of vagal stimulation on the overflow of
norepinephrine into the coronary sinus during cardiac sympathetic ner
ve stimulation in the dog.-Circ. Res.. 1976, 38: 81-85.
24. Watanabe A. M., Besch H. R. Interaction between cyclic adenosine mo-
nophosphate and cyclic guanosine monophosphate in guinea pig ventri
cular myocardium.-Circ. Res., 1975, 37: 309-317.
25. Surawicz B. Ventricular fibrillation.-Am. J. Cardiol., 1971
26. Petropoulos P. C, Jaijne N. G. Cardiac function during perfusion of the
circumflex coronary artery with venous blood, low molecular weignt
dextran in Tyrode solution.-Am. Heart J., 1964, 68: 370-382.
27. Sewell W. M., Koth D. R., Huggins С . Е . Ventricular fibrillation in dogs
after sudden return of flow to the coronary artery.-Surgery, 1955, 38
1050-1053.
28. Bagdonas A. A., Stuckey J. H., Piera J. Effects of ischemia and hypoxia
on the specialized conducting system of the canine heart.-Am. Heart
J., 1961, 61: 206-218.
29. Danese С Pathogenesis of ventricular fibrillation in coronary occlusion.-
JAMA, 1962, 179: 52-53.
30. Kerzner J., Wolf U., Kosowsky B. D. et al. Ventricular ectopic rhythms
following vagal stimulation in dogs with acute myocardial infarction.-
Circulation, 1973, 47:44-50.
31. Haggins С . В ., Vainer S. F., Braunwald E. Parasympathetic control of
the heart.-Pharmacol. Rev., 1973, 25: 119-155.
32. Verrier R. L., Lown B. Effect of left stellectomy on enhanced cardiac
vulnerability induced by psychologic stress (abstr.).-Circulation, 1977,
56:111-80.
33. Nathanson M. H. Action of acetyl beta methyolcholin on ventricular
hrythm induced by adrenalin.-Proc. Soc. Exp. Biol. Med., 1935, 32: 1297-1299.
34. Cope R. L. Suppressive effect of carotid sinus on premature ventricular
beats in certain instances.-Am. J. Cardiol., 1959, 4: 314-320.
35. Lown В ., Levine S. A. The carotid sinus: clinical value of its stimulati
on.-Circulation, 1961, 23: 776-789.
36. Lorentzen D. Pacemaker-induced ventricular tacycardia: reversion to
normal sinus rhythm by carotid sinus massage.-JAMA, 1976, 235: 282-283.
37. Waxman M. В ., Downar E., Berman D. et al. Phenylephrine (Neosyne-
phrine R) terminated ventricular tachycardia.-Circulation, 1974, 50:
38. Weiss Т ., Lattin G. M., Engelman K. Vagally mediated suppression of
premature ventricular contractions in man.-Am. Heart J., 1975, 89: 700-707.
39. Lown В ., Graboys Т . В ., Podrid P. J. et al. Effect of a digitalis drug on
ventricular premature beats (VPBs).-N. Engl. J. Med., 1977, 296: 301-306.
Оглавление темы "Механизмы регуляции деятельности сердца. Венозный возврат крови к сердцу. Центральное венозное давление (ЦВД). Параметры гемодинамики.":2. Механизмы регуляции деятельности сердца. Адренергические механизмы регуляции сердца.
3. Холинергические механизмы регуляции сердца. Влияние ацетилхолина на сердце.
4. Рефлекторные влияния на сердце. Кардиальные рефлексы. Рефлекс Бейнбриджа. Рефлекс Генри-Гауэра. Рефлекс Данини-Ашнера.
5. Гуморальные (гормональные) влияния на сердце. Гормональная функция сердца.
6. Венозный возврат крови к сердцу. Величина венозной крови притекающей к сердцу. Факторы влияющие на венозный возврат.
7. Уменьшение венозного возврата. Увеличение венозного возврата крови к сердцу. Спланхническое сосудистое русло.
8. Центральное венозное давление (ЦВД). Величина центрального венозного давления (ЦВД). Регуляция цвд.
9. Параметры гемодинамики. Соотношение основных параметров системной гемодинамики.
10. Регуляция сердечного выброса. Изменение оцк. Компенсаторные реакции сосудистой системы.
Влияние симпатических нервов на сердце проявлется в виде положительного хронотропного и положительного инотропного эффекта. Сведения о наличии тонических влияний симпатической нервной системы на миокард основываются в основном на хронотропных эффектах.
Электрическая стимуляция волокон, отходящих от звездчатого ганглия, вызывает увеличение ритма сердца и силы сокращений миокарда (см. рис. 9.17). Под влиянием возбуждения симпатических нервов скорость медленной диастолической деполяризации повышается, снижается критический уровень деполяризации клеток водителей ритма синоатриального узла, уменьшается величина мембранного потенциала покоя. Подобные изменения увеличивают скорость возникновения потенциала действия в клетках водителей ритма сердца, повышают его возбудимость и проводимость. Эти изменения электрической активности связаны с тем, что выделяющийся из окончаний симпатических волокон медиатор норадреналин взаимодействует с B1-адренорецепторами поверхностной мембраны клеток, что приводит к повышению проницаемости мембран для ионов натрия и кальция, а также уменьшению проницаемости для ионов калия.
Рис. 9.17. Электрическое раздражение эфферентных нервов сердца
Ускорение медленной спонтанной диастолической деполяризации клеток водителей ритма, увеличение скорости проведения в предсердиях, атриовентрикулярном узле и желудочках приводит к улучшению синхронности возбуждения и сокращения мышечных волокон и к увеличению силы сокращения миокарда желудочков. Положительный инотропный эффект связан также с повышением проницаемости мембраны для ионов кальция. При увеличении входящего тока кальция возрастает степень электромеханического сопряжения, в результате чего увеличивается сократимость миокарда.

Менее изучено участие в регуляции сердечной деятельности интракардиальных ганглиозных нервных элементов. Известно, что они обеспечивают передачу возбуждения с волокон блуждающего нерва на клетки синоат-риального и атриовентрикулярного узлов, выполняя функцию парасимпатических ганглиев. Описаны инотропные, хронотропные и дромотропные эффекты, полученные при стимуляции этих образований в условиях эксперимента на изолированном сердце. Значение этих эффектов в естественных условиях остается неясным.
Оглавление темы "Возбудимость сердечной мышцы. Сердечный цикл и его фазовая структура. Тоны сердца. Иннервация сердца.":1. Возбудимость сердечной мышцы. Потенциал действия миокарда. Сокращение миокарда.
2. Возбуждение миокарда. Сокращение миокарда. Сопряжение возбуждения и сокращения миокарда.
3. Сердечный цикл и его фазовая структура. Систола. Диастола. Фаза асинхронного сокращения. Фаза изометрического сокращения.
4. Диастолический период желудочков сердца. Период расслабления. Период наполнения. Преднагрузка сердца. Закон Франка-Старлинга.
5. Деятельность сердца. Кардиограмма. Механокардиограмма. Электрокардиограмма (ЭКГ). Электроды экг.
6. Тоны сердца. Первый (систолический) тон сердца. Второй (диастолический) сердечный тон. Фонокардиограмма.
7. Сфигмография. Флебография. Анакрота. Катакрота. Флебограмма.
8. Сердечный выброс. Регуляция сердечного цикла. Миогенные механизмы регуляции деятельности сердца. Эффект Франка - Старлинга.
9. Иннервация сердца. Хронотропный эффект. Дромотропный эффект. Инотропный эффект. Батмотропный эффект.
Результатом стимуляции этих нервов является отрицательный хронотропный эффект сердца (рис. 9.17), на фоне которого проявляются также отрицательные и дромотропный инотропный эффекты . Существуют постоянные тонические влияния на сердце со стороны бульбарных ядер блуждающего нерва: при его двусторонней перерезке частота сердцебиений возрастает 1,5-2,5 раза. При длительном сильном раздражении влияние блуждающих нервов на сердце постепенно ослабевает или прекращается, что получило название «эффекта ускользания» сердца из-под влияния блуждающего нерва.
Различные отделы сердца по-разному реагируют на возбуждение парасимпатических нервов . Так, холинергические влияния на предсердия вызывают значительное угнетение автоматии клеток синусного узла и спонтанно возбудимой ткани предсердий. Сократимость рабочего миокарда предсердий в ответ на стимуляцию блуждающего нерва снижается. Рефрактерный период предсердий при этом также уменьшается в результате значительного укорочения длительности потенциала действия предсердных кардиомиоцитов. С другой стороны, рефрактерность кардиомиоцитов желудочков под влиянием блуждающего нерва, напротив, значительно возрастает, а отрицательный парасимпатический инотропный эффект на желудочки выражен в меньшей степени, чем на предсердия.
Рис. 9.17. Электрическое раздражение эфферентных нервов сердца . Вверху - уменьшение частоты сокращений при раздражении блуждающего нерва; внизу-увеличение частоты и силы сокращений при раздражении симпатического нерва. Стрелками отмечены начало и конец раздражения.Электрическая стимуляция блуждающего нерва вызывает урежение или прекращение сердечной деятельности вследствие торможения автоматической функции водителей ритма синоатриального узла. Выраженность этого эффекта зависит от силы и частоты . По мере увеличения силы раздражения отмечается переход от небольшого замедления синусного ритма до полной остановки сердца.
Отрицательный хронотропный эффект раздражения блуждающего нерва связан с угнетением (замедлением) генерации импульсов в водителе ритма сердца синусного узла. Поскольку при раздражении блуждающего нерва в его окончаниях выделяется медиатор - ацетилхолин , при его взаимодействии с мускариночувствительными рецепторами сердца повышается проницаемость поверхностной мембраны клеток водителей ритма для ионов калия. Вследствие этого возникает гиперполяризация мембраны, которая замедляет (подавляет) развитие медленной спонтанной диастолической деполяризации, и поэтому мембранный потенциал позже достигает критического уровня. Это приводит к урежению ритма сокращений сердца.
При сильных раздражениях блуждающего нерва диастолическая деполяризация подавляется, возникают гиперполяризация водителя ритма и полная остановка сердца. Развитие гиперполяризации в клетках водителей ритма снижает их возбудимость, затрудняет возникновение очередного автоматического потенциала действия и тем самым приводит к замедлению или даже остановке сердца. Стимуляция блуждающего нерва , усиливая выход калия из клетки, увеличивает мембранный потенциал, ускоряет процесс реполяризации и при достаточной силе раздражающего тока укорачивает длительность потенциала действия клеток водителя ритма.
При вагусных воздействиях имеет место уменьшение амплитуды и длительности потенциала действия кардиомиоцитов предсердия. Отрицательный инотропный эффект связан с тем, что уменьшенный по амплитуде и укороченный потенциал действия не способен возбудить достаточное количество кардиомиоцитов. Кроме того, вызванное ацетилхолином повышение калиевой проводимости противодействует потенциалзависимому входящему току кальция и проникновению его ионов внутрь кардиомиоцита. Холинергический медиатор ацетилхолин может также угнетать АТФ-азную активность миозина и, таким образом, уменьшать величину сократимости кардиомиоцитов. Возбуждение блуждающего нерва приводит к повышению порога раздражения предсердий, подавлению автоматии и замедлению проводимости атриовентрикулярного узла. Указанное замедление проводимости при холинергических влияниях может вызвать частичную или полную атриовентрикулярную блокаду.
Учебное видео иннервации сердца (нервов сердца)
При проблемах с просмотром скачайте видео со страницыНервная регуляция работы сердца осуществляется симпатическими и парасимпатическими импульсами. Первые повышают частоту, силу сокращений, давление крови, а вторые имеют противоположное влияние. Возрастные изменения тонуса вегетативной нервной системы учитывают при назначении лечения.
📌 Читайте в этой статье
Особенности симпатической нервной системы
Симпатическая нервная система предназначена для активизации всех функций организма при стрессовой ситуации. Она обеспечивает реакцию по типу «сражайся или беги». Под влиянием раздражения нервных волокон, которые в нее входят, происходят такие изменения:
- слабый бронхоспазм;
- сужение артерий, артериол, особенно расположенных в коже, кишечнике и почках;
- сокращение матки, сфинктеров мочевого пузыря, капсулы селезенки;
- спазм радужной мышцы, расширение зрачка;
- понижение двигательной активности и тонуса кишечной стенки;
- ускоренная .
Усиление всех сердечных функций – возбудимости, проводимости, сократимости, автоматизма, расщепление жировой ткани и выброс почками ренина (повышает давление) связаны с раздражением бета-1 адренорецепторов. А стимуляция бета-2 типа приводит к:
- расширению бронхов;
- расслаблению мышечной стенки артериол в печени и мышцах;
- расщеплению гликогена;
- выбросу инсулина для проведения глюкозы внутрь клеток;
- образованию энергии;
- снижению маточного тонуса.
Симпатическая система не всегда оказывает однонаправленное действие на органы, что связано с наличием в них нескольких видов адренорецепторов. В конечном итоге в организме возрастает переносимость физических и умственных нагрузок, происходит усиление работы сердца и скелетной мускулатуры, перераспределение кровообращения для питания жизненно важных органов.
Чем отличается парасимпатическая система
Этот отдел вегетативной нервной системы предназначен для расслабления организма, восстановления после нагрузки, обеспечения пищеварения и накопления энергии. При активизации блуждающего нерва:
- возрастает приток крови к желудку и кишечнику;
- увеличивается выброс пищеварительных ферментов и продукция желчи;
- сужаются бронхи (в покое не требуется много кислорода);
- замедляется ритм сокращений, падает их сила;
- снижается тонус артерий и .
Влияние двух систем на сердце
Несмотря на то, что на сердечно-сосудистую систему симпатическая и парасимпатическая стимуляция оказывают противоположное воздействие, это не всегда так однозначно проявляется. А механизмы их взаимовлияния не имеют математической закономерности, не все они достаточно изучены, но установлено:
- чем больше повышается симпатический тонус, тем сильнее будет подавляющий эффект парасимпатического отдела – акцентированное противодействие;
- при достижении нужного результата (например, ускорения ритма при нагрузке) тормозится симпатическое и парасимпатическое влияние – функциональный синергизм (однонаправленное действие);
- чем выше начальный уровень активации, тем меньше возможность его возрастания при раздражении – закон исходного уровня.
Смотрите на видео о влиянии на сердце симпатической и парасимпатической систем:
Влияние возраста на вегетативный тонус
У новорожденных преобладает влияние симпатического отдела на фоне общей незрелости нервной регуляции. Поэтому у них существенно ускорен. Затем обе части вегетативной системы развиваются очень быстро, достигая максимума к подростковому периоду. В это время отмечается наивысшая концентрация нервных сплетений в миокарде, что объясняет быструю смену давления и скорости сокращений при внешних воздействиях.
До 40 лет преобладает парасимпатический тонус, что сказывается на замедлении пульса в состоянии покоя и быстром возвращением его к норме после нагрузок. А затем начинаются возрастные изменения – сокращается количество адренорецепторов при сохранении парасимпатических ганглиев. Это приводит к следующим процессам:
- ухудшается возбудимость мышечных волокон;
- нарушаются процессы образования импульсов;
- повышается чувствительность сосудистой стенки и миокарда к действию гормонов стресса.
Под воздействием ишемии клетки приобретают еще большую реакцию на симпатические импульсы и реагируют даже на малейшие сигналы спазмом артерий и ускорением пульса. При этом возрастает электрическая нестабильность миокарда, что объясняет частое появление при , а особенно при .
Доказано, что нарушения симпатической иннервации во много раз превышают зону разрушения при остром нарушении коронарного кровообращения.
Что происходит при возбуждении
В сердце находятся в основном бета 1 адренорецепторы, немного бета 2 и альфа типа. При этом они расположены на поверхности кардиомиоцитов, что повышает их доступность для основного медиатора (проводника) симпатических импульсов – норадреналина. Под влиянием активизации рецепторов происходят такие изменения:
- повышается возбудимость клеток синусового узла, проводящей системы, мышечных волокон, они реагируют даже на подпороговые сигналы;
- ускоряется проведение электрического импульса;
- возрастает амплитуда сокращений;
- увеличивается количество ударов пульса за минуту.
На наружной мембране клеток сердца обнаружены и парасимпатические холинорецепторы типа М. Их возбуждение тормозит активность синусового узла, но одновременно повышает возбудимость предсердных мышечных волокон. Этим можно объяснить развитие наджелудочковой экстрасистолии ночью, когда высок тонус блуждающего нерва.
Вторым депрессивным эффектом является угнетение парасимпатической системой проведения в атриовентрикулярном узле, что задерживает распространение сигналов на желудочки.
Таким образом, парасимпатическая нервная система:
- снижает возбудимость желудочков и повышает ее в предсердиях;
- замедляет сердечный ритм;
- тормозит образование и проведение импульсов;
- подавляет сократительную способность мышечных волокон;
- уменьшает потребность миокарда в кислороде;
- препятствует спазму стенок артерий и .
Симпатикотония и ваготония
В зависимости от преобладания тонуса одного из отделов вегетативной нервной системы у пациентов может быть исходное повышение симпатических влияний на сердце – симпатикотония и ваготония при избыточной активности парасимпатических. Это имеет значение при назначении лечения заболеваний, так как реакция на медикаменты может быть разная.
Например, при исходной симпатикотонии у пациентов можно выявить:
- кожа сухая и бледная, конечности холодные;
- пульс ускорен, преобладает повышение систолического и пульсового давления;
- сон нарушен;
- психологически устойчивы, активны, но есть высокая тревожность.
Для таких больных нужно использовать успокаивающие препараты и адреноблокаторы как основу медикаментозной терапии. При ваготонии кожа влажная, есть склонность к обморокам при резкой перемене положения тела, движения замедлены, переносимость нагрузок низкая, разница систолического и диастолического давления уменьшена.
Для терапии целесообразно использовать антагонисты кальция, .
Симпатические нервные волокна и медиатор норадреналин обеспечивают активность организма при действии стрессовых факторов. При стимуляции адренорецепторов повышается давление, ускоряется пульс, повышается возбудимость и проводимость миокарда.
Парасимпатический отдел и ацетилхолин имеют противоположную направленность влияния на сердце, они отвечают за расслабление и накопление энергии. В норме эти процессы последовательно сменяют друг друга, а при нарушении нервной регуляции (симпатикотония или ваготония) изменяются показатели кровообращения.
Читайте также
Существуют гормоны сердца. Они оказывают влияние на работу органа - усиливающую, замедляющую. Это могут быть гормоны надпочечников, щитовидной железы и другие.