Схема строения клеток эукариот и прокариот. Строение прокариотической (бактериальной) клетки
Читайте также
Для всех микроорганизмов, входящих в царство, характерен прокариотический тип организации клетки, что определяется особенностями их ультраструктуры, а также строения и функций ряда макромолекул. Из всех известных клеток прокариотическая самая простая и, вероятно, первая клетка, возникшая около 3,6 млрд. лет назад.
В настоящее время предполагают, что в какой-то момент времени эволюция клеток пошла по двум самостоятельным направлениям. Появились две группы организмов -прокариоты, у которых ядерный материал не был ограничен оболочкой, и эукариоты, имеющие оформленное ядро с ядерной оболочкой.
Основные отличия прокариотов от эукариотов состоят в следующем:
в клетках прокариотов отсутствуют компартменты, или органеллы, ограниченные от цитоплазмы специализированными внутриклеточными липопротеидными мембранами: эндоплазматическая сеть (ретикулюм), митохондрии, аппарат Гольджи, лизосомы, хлоропласты;
ядерная структура прокариотов, называемая нуклеоидом, не имеет ядерной оболочки с поровым комплексом и представлена макромолекулой ДНК с белками (без гистонов);
геном прокариотической клетки организован в одну кольцевую хромосому, которая представляет собой единый репликон и не делится митозом;
дополнительные репликоны могут быть представлены кольцевыми молекулами плазмидных ДНК;
прокариотическая клетка содержит только один тип рибосом с константой седиментации 708, причем часть рибосом ассоциирована с цитоплазматической мембраной, что никогда не наблюдается у эукариотов;
клеточная стенка прокариотов содержит характерный только для бактерий биогетерополимер - пептидогликан.
Некоторые прокариоты обладают структурами, отсутствующими у эукариотов:
подвижные бактерии имеют особые бактериальные жгутики из белков-флагеллинов;
спорообразующие формы бактерий в неблагоприятных условиях превращаются в уникальные по степени устойчивости типы покоящихся клеток - бактериальные споры;
клетки прокариотов очень малы; диаметр большинства клеток бактерий не превышает 1 мкм, однако длина может быть значительной, например, у некоторых спирохет - до 500 мкм. Малые размеры прокариотов связаны, как полагают, с отсутствием в их ультраструктуре специализированных мембранных систем, что затрудняет координацию внутриклеточных процессов пропорционально увеличению размеров клетки.
Клеточное строение четко отделяет прокариотов от вирусов. Подчеркивая примитивность организации бактериальных клеток, необходимо однако отметить, что они эволюционировали в своем направлении в течение гораздо большего времени, чем эукарио-тические, и, хотя эволюционные возможности прокариотической клетки, по всей видимости, ограничены, в процессе эволюции происходили изменения их клеточной организации, что привело постепенно к ее усложнению.
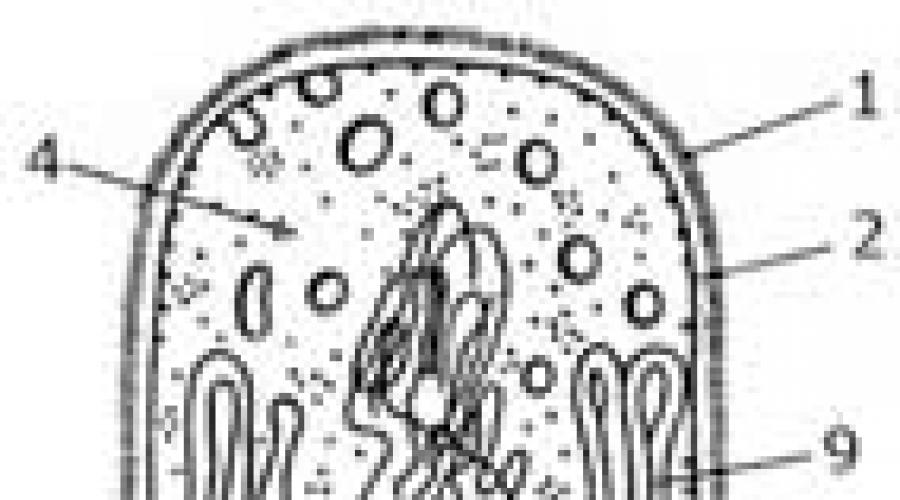
По целому ряду признаков бактерии имеют принципиальные различия с эукариотами, и знание особенностей их устройства и функционирования позволяет понять возможность избирательного антимикробного действия химиотерапевтических препаратов. Применение электронной микроскопии и тонких цитохимических исследований позволило изучить их ультраструктуру (рис. 1). Обязательными компонентами бактериальной клетки являются цитоплазматическая мембрана, окружающая цитоплазму, в которой содержатся рибосомы и нуклеоид. Клетки всех бактерий, за исключением L-форм и микоплазм, имеют клеточную стенку. Другие структуры являются дополнительными и определяют морфологические и функциональные особенности различных видов: капсулы, жгутики, пили, споры, включения.
Рис. 1. Схема строения прокариотической клетки:
/ - капсула; 2 - клеточная стенка; 3 - цитоплазматическая мембрана; 4 - мезосомы; 5 - цитоплазма; 6 - нуклеоид; 7 - плазмида;
8 - рибосомы и полисомы; 9 - жгутики; 10 - пили; 11 - гранулы гликогена; 12 - липидные капельки; 13 - гранулы волютина; 14 - включения серы
Поверхностные структуры. Капсула - это наружный, самый верхний слизистый слой клетки различной толщины фибриллярной или глобулярной структуры. Она имеет полисахаридную, му-кополисахаридную или полипептидную природу и содержит до 98 % воды. В зависимости от толщины различают микрокапсулу (толщиной менее 0,2 мкм) и макрокапсулу. Капсула не является обязательным структурным элементом клетки. Биологический смысл капсулообразования определяется целым рядом функций, среди которых: защита от фагоцитов и вирусов, токсинов и радиации; иммунологическая мимикрия у патогенных бактерий; сохранение влаги в условиях пониженной влажности; прикрепление клетки к плотной поверхности.
Пили (фимбрии, ворсинки, реснички) - это прямые цилиндрические образования белковой природы длиной 0,3-10 мкм, диаметром до 10 нм, равномерно покрывающие поверхность клетки (до нескольких сотен на клетку), не выполняющие локомоторную функцию.
Различают пили общего типа, способствующие прикреплению бактериальной клетки к субстрату, клеткам человека (явление адгезии микроорганизмов) и пили половые, участвующие в передаче генетического материала от клетки-донора к клетке-реципиенту в процессе конъюгации, а также обуславливающие адсорбцию специфических бактериофагов на клетках.
Жгутики - органы движения бактерий в виде спирально изогнутых цилиндрических образований белковой природы (белки-флагеллины) на поверхности клетки длиной 3-12 мкм и толщиной 10-30 нм, прикрепленные базальным телом (системой дисков) к цитоплазматической мембране (см. вкл. I). Число и располо-
жение жгутиков может быть различным и является видовым признаком (рис. 2). Различают монотрихи (бактерии с одним жгутиком на конце), амфитрихи (бактерии с жгутиками, расположенными по полюсам), лофотрихи (клетки с пучком жгутиков на одном конце) и перитрихи (с 2-30 жгутиками по всему телу клетки).
Пили и жгутики не являются обязательными органоидами бактериальной клетки.
Клеточная стенка - один из основных структурных элементов бактерии, выполняющий механическую защиту клетки. Кроме микоплазм и L-форм, клетки всех бактерий покрыты клеточной стенкой, толщина которой у разных видов колеблется в пределах 0,01-14 мкм. Она представляет собой плотную эластичную структу -
Рис. 2. Основные формы бактерий (по А. А. Воробьеву и соавт., 1994):
/ - стафилококки; 2 - стрептококки; 3 - сарцины; 4 - гонококки;
5- пневмококки; 6- капсулы пневмококков; 7- коринебактерии дифтерии; 8 - клостридии; 9 - бациллы; 10 - вибрионы; 11 - спириллы; 12 - трепонсмы; - боррелии; 14 - лептоспиры; 15- актиномицеты; 16 - расположение жгутиков: а - монотрихи; б - лофотрихи; в - амфитрихи; г - перитрихи
ру, которая окружает протопласт клетки и придает ей постоянную форму и жесткость. Клеточная стенка препятствует осмотическому набуханию и разрыву клеток, когда они попадают в гипотоническую среду. Вода, другие малые молекулы и разные ионы легко проникают через крошечные поры в клеточной стенке, но через них не проходят крупные молекулы белков и нуклеиновых кислот.
Основным химическим компонентом клеточной стенки является специфический гетерополимер - пептидогликан (муреин, мукопептид, глюкозаминопептид, гликопептид), состоящий из цепочек, в которых чередуются остатки N-ацетилглюкозамина и М-ацетилмурамовой кислоты, соединенные между собой β-1,4-гликозидными связями. Это резко отличает оболочечные структуры бактерий от эукариотических и создает «ахиллесову пяту» бактерий, используемую для антимикробной химиотерапии.
Организация цитоплазмы. Цитоплазматическая мембрана (ЦМ) Носится к числу обязательных клеточных структур, имеет толщину 7-13 нм и располагается непосредственно под клеточной стенкой, ограничивая протопласт клетки. По своему строению мембраны бактериальных, животных и растительных клеток очень сходны. В настоящее время большинством ученых принята жидкостно-мозаичная модель строения ЦМ. Согласно этой модели ЦМ состоит из двойного слоя (15-30 % молекул фосфолипидов и триглицеридов с направленными внутрь гидрофобными концами и гидрофильными «головками» наружу. В него мозаично погружены молекулы белка (50-70%). В мембране имеются также углеводы (2-5 %) и РНК. ЦМ представляет собой пластическое «текучее» образование, которое играет важнейшую роль обмене веществ, является полупроницаемой структурой, поддерживает осмотическое давление, контролирует как поступление веществ в клетку, так и выведение конечных метаболитов по системе субстрат специфичных пермеаз (ферментов-переносчиков, локализованных на мембране). С ЦМ связаны процессы дыхания, доставляющие клетке энергию, то есть те функции, за которые в эукариотической клетке ответственны мембраны митохондрий и хлоропластов.
Выделяют так называемые мезосомы - впячивания ЦМ - смешанные мембранные системы, образованные трубочками, пузырьками и ламеллами. Предполагается выполнение ими функций центра дыхательной активности бактерий, участие в делении клетки и расхождении дочерних хромосом после репликации.
Цитоплазма заполняет собой объем бактерии, ограниченный ЦМ. Это сложная коллоидная система, которая состоит из бельков, нуклеиновых кислот, углеводов, липидов, минеральных веществ и 70-80 % воды. Цитоплазма является местонахождением внутриклеточных органелл (нуклеоида, рибосом, различных включений) и участвует во внутриклеточном метаболизме. Характер-
ными чертами организации цитоплазмы прокариотов по сравнению с животными и растительными клетками являются отсутствие эндоплазматического ретикулюма и высокая электронная плотность.
Нуклеоид - ядерный материал бактериальной клетки. Он представлен двойной нитью макромолекулы ДНК с молекулярной массой 2-3 10 в соединении с белками, среди которых отсутствуют характерные для эукариотов ядерные (гистоны и гистоноподобные) белки. В отличие от настоящего ядра эукариотических клеток нуклеоид не имеет ядерной перфорированной мембраны, не делится митозом и представляет собой в период деления одну кольцевую хромосому, кодирующую всю генетическую информацию.
Плазмиды - необязательные внутриклеточные структуры в виде внехромосомных кольцевых участков ДНК, способных к саморепликации. Обуславливают наследование дополнительных признаков: лекарственной устойчивости, токсигенности, бактериоциногенности и др.
Рибосомы - органеллы, в которых осуществляется синтез белка. Каждая рибосома имеет размеры 20х30х30 нм и константу седиментации 70S (так как при ультрацентрифугировании рибосомы оседают со скоростью около 70 единиц Сведенберга (S), в отличие от более крупных цитоплазматических рибосом эукариотов с константой седиментации 808). В свободном состоянии бактериальная рибосома находится в виде двух субъединиц - 30S и 50S, обе субъединицы содержат примерно по 40 % рибосомальной РНК и 60 % белка. Во время синтеза белка рибосомы с помощью информационной РНК образуют полисомы, обычно связанные с ЦМ. Бактерии могут содержать от 5000 до 50 000 рибосом, что зависит от возраста клетки и условий культивирования.
Знание различий между рибосомами бактерий и эукариотических клеток имеет важное значение для понимания механизмов антимикробного действия тех антибиотиков, которые подавляют синтез белка на бактериальных рибосомах и не затрагивают функции 80S рибосом.
Споры (эндоспоры) бактерий - покоящиеся формы некоторых видов грамположительных бактерий в неблагоприятных условиях внешней среды.
Спорообразование происходит в несколько стадий, при полном созревании споры вегетативная часть клетки лизируется и отмирает (см. вкл. I, II).
В процессе спорообразования (споруляции) можно выделить несколько основных этапов. Переходящая к спорообразованию клетка перестает расти; как правило, она содержит два и более нуклеоидов. На первом этапе часть клеточной ДНК локализуется в одном из полюсов клетки. Затем часть цитоплазмы с заключен-
Иной в ней хромосомой обособляется цитоплазматической мембраной, как бы врастающей в глубь клетки, при этом образуется проспорь, окруженная двойной мембранной оболочкой.
Затем между двумя мембранами идет формирование многослойной стенки и коры (кортекса) споры пептидогликановой природы. Снаружи мембран образуется также полипептидная оболочка и экзоспорий, окружающий спору в виде свободного чехла. Полностью сформировавшаяся бактериальная спора - это уплотненный участок клетки с нуклеоидом и рибосомами, ограниченный плотной многослойной оболочкой, пропитанной кальциевыми Солями дипиколиновой кислоты.
Спорообразование характерно для палочковидных бактерий - бацилл и клостридий (см. рис. 2). Различают центральное, терминальное и субтерминальное расположение спор в вегетативной части клетки, что является дифференциально-диагностическим признаком возбудителя.
В одной бактерии образуется одна спора, находящаяся в стадии покоя, при этом все процессы обмена веществ практически сведены к нулю, но сохраняется потенциальная жизнеспособность клетки. Поскольку увеличение числа микроорганизмов в этом процессе не происходит, спорообразование у бактерий не является способом размножения, а лишь приспособлением для выживания. Уникальные по степени своей устойчивости к физическим и химическим факторам бактериальные споры могут сохраняться до внешней среде без потери жизнеспособности длительное время (десятки лет), затрудняя борьбу со спороносными патогенными бактериями.
Внутриплазматические включения. Термином «включения» обозначают такие внутриклеточные структуры бактерий, которые, очевидно, не являются абсолютно необходимыми для их жизнедеятельности. Однако их природа и функции могут быть различны. В одних случаях включения являются продуктами обмена бактериальной клетки, в других - запасом питательных веществ.
Из резервных полисахаридов особенно распространены глюканы - гликоген, крахмал, гранулеза. Они выявляются в клетках бацилл, клостридий, энтеробактерий и др.
Запасные липиды представлены полиэфиром β-оксимасляной кислоты и восками. Воски, эфиры высокомолекулярных жирных кислот и спиртов характерны для микобактерий.
У коринебактерий резерв фосфора создается в виде зерен полифосфатов (волютина), имеющих диагностическое значение.
В клетке. Наследственная информация передается с помощью кольцевой ДНК , которая прикреплена к плазматической оболочке клетки . Кроме этого, в клетках прокариотов присутствуют рибосомы, а цитоплазма имеет гелевую консистенцию, которая обеспечивает устойчивость к высоким температурам. Размножаются прокариоты с помощью простейшего деления, без полового процесса. Многоклеточных форм у таких организмов не бывает.
Строение прокариотической и эукариотической клетки. Различия в строении.
Царство Дробянки - это упраздненное сегодня царство живых организмов, которым раньше называли прокариотов. Название образовано от способа размножения прокариотов - делением. Прокариоты появились на земле более 3,5 млрд.лет назад.
Подцарство Архебактерии - это самые древние прокариоты, которые отличаются от других видов прокариотов строением и отсутствием муреина (пептидогликана) в клеточных стенках бактерий. Кольцевая ДНК архебактерий построена по типу эукариотических организмов - по типу избыточного генома.
Архебактерии делятся на три типа :
1) Галобактерии ;
2) Метаногенные бактерии ;
3) Экстремальные термофилы .
Галобактерии практикуют фотосинтез без выделения кислорода с пигментом бактериородопсином.
Эубактерии - это самая многочисленная группа микроорганизмов, клеточная оболочка которых имеет муреин в своем составе. Делятся на граммотрицательные бактерии и граммположительные бактерии (определяют с помощью реакции на анилиновые красители). Эубактерии могут образовывать споры, а размножаются они с помощью некоего подобия полового процесса - конъюгацией , обмениваясь плазмидами. Плазмиды - это небольшые кольцевые ДНК, внехромосомные частицы, которые содержат не более одного гена.
По форме клеток различают следующие виды эубактерий:
- Бактерии;
- Кокки;
- Вибрионы;
- Бациллы;
- Спирохеты;
- Спириллы.
По типу питания эубактерии бывают фотоавтотрофами (без выделения кислорода), хемотрофами и гетеротрофами . Бывают аэробные бактерии и анаэробные бактерии .
Эубактерии играют важную роль в общей биосистеме:
1) Выполняют геологическую роль (железобактерии , серные бактерии и др.);
2) Принимают непосредственное участие в круговороте веществ (сапротрофы );
3) Являются возбудителями различных заболеваний у других живых организмов, в том числе и людей;
4) Используются человеком для своих целей - в виноделии, сыроделании, образовании аминокислот, кормового белка, витаминов и др.
Подцарство Оксифотобактерии . Это подцарство делится на два отдела: хлороксибактерии и цианобактерии (сине-зеленые водоросли). К хлороксибактериям относятся прокариоты рода прохлорон, которые были открыты во второй половине 20-го века. Ученые до сих пор дискутируют насчет их происхождения. Они обитают в симбиозе с асцидиями в морях тропиков и субтропиков. Их набор фотосинтетических элементов является таким же, как у зеленых водорослей и высших растений.
Прокариотические клетки - это наиболее примитивные, очень просто устроенные, сохраняющие черты глубокой древности организмы. К прокариотическим (или доядерным) организмам относят бактерии и синезеленые водоросли (цианобактерии). На основании общности строения и резких отличий от других клеток прокариотические выделяют в самостоятельное царство дробянки.
Рассмотрим строение прокариотической клетки на примере бактерий. Генетический аппарат прокариотической клетки представлен ДНК единственной кольцевой хромосомы , находится в цитоплазме и не отграничен от нее оболочкой. Такой аналог ядра называют нуклеоидом. ДНК не образует комплексов с белками и поэтому все гены, входящие в состав хромосомы, "работают", т.е. с них непрерывно считывается информация.
Прокариотическая клетка окружена мембраной, отделяющей цитоплазму от клеточной стенки, образованной из сложного, высокополимерного вещества. В цитоплазме органелл мало, но присутствуют многочисленные мелкие рибосомы (бактериальные клетки содержат от 5000 до 50 000 рибосом).
 Цитоплазма прокариотической клетки пронизана мембранами, образующими эндоплазматическую сеть, в ней и находятся рибосомы, осуществляющие синтез белков.
Цитоплазма прокариотической клетки пронизана мембранами, образующими эндоплазматическую сеть, в ней и находятся рибосомы, осуществляющие синтез белков.
Внутренняя часть клеточной стенки прокариотической клетки представлена плазматической мембраной, выпячивания которой в цитоплазму образуют мезосомы, участвующие в построении клеточных перегородок, репродукции, и являются местом прикрепления ДНК. Дыхание у бактерий осуществляется в мезосомах, у сине-зеленых водорослей в цитоплазматических мембранах.
У многих бактерий внутри клетки откладываются запасные вещества: полисахариды, жиры, полифосфаты. Резервные вещества, включаясь в обмен веществ, могут продлевать жизнь клетки в отсутствие внешних источников энергии.
(1-клеточная стенка, 2-наружная цитоплазматическая мембрана, 3-хромосома(кольцевая молекула ДНК), 4-рибосома, 5-мезосома, 6-впячивание наружной цитоплазмотической мембраны, 7-вакуоли, 8-жгутики, 9-стопки мембран, в которых осуществляется фотосинтез)
Как правило, бактерии размножаются делением надвое. После удлинения клетки постепенно образуется поперечная перегородка, закладывающаяся в направлении снаружи внутрь, затем дочерние клетки расходятся или остаются связанными в характерные группы - цепочки, пакеты и т.д. Бактерия - кишечная палочка каждые 20 минут удваивает свою численность.
Для бактерий характерно спорообразование. Оно начинается с отшнуровывания части цитоплазмы от материнской клетки. Отшнуровавшаяся часть содержит один геном и окружена цитоплазматической мембраной. Затем вокруг споры вырастает клеточная стенка, нередко многослойная. У бактерий наблюдается половой процесс в форме обмена генетической информацией между двумя клетками. Половой процесс повышает наследственную изменчивость микроорганизмов.
Большинство живых организмов объединено в надцарство эукариот, включающих царство растений, грибов и животных. Эукариотические клетки крупнее прокариотических клеток , состоят из поверхностного аппарата, ядра и цитоплазмы.
Немного истории. Аристотель пытался все объекты природы систематизировать. У него была "лестница существ". Внизу наиболее примитивно организованные камни, потом растения, животные и человек. Стремление к линейной классификации довольно долго сохранялось, но потом его пришлось отвергнуть, так как объекты живой природы не выстраиваются в единую лестницу. Деление на растения и животные известно давно. Эти группы называют царствами: царство растений и царство животных. Потом были описаны простые одноклеточные растения и животные, о которых не всегда понятно, растения они или животные. Они были выделены в группу одноклеточные (Протисты). Затем открыли бактероий и выделили их в отдельное царство. Позже было в отдельное царство были выделены грибы. Для нас они кажутся похожими на растения, но, тем не менее, от растений они существенно отличаются, в частности, тем, что, как животные, запасают гликоген, а не крахмал.
Итак, живые организмы разделили на царства Растений, Грибов, Животных и Простейших (одноклеточных), и царство бактерий, в которую входили все прокариоты. Но когда изучали бактерий, оказалось, что они также делятся на две сильно отличающиеся группы. Соответственно, их пришлось разделить на два царства: Эубактерии (собственно бактерии) и Архебактерии (другое название – Археи). Последние также не имеют ядра, но по структуре сильно отличаются от бактерий.
Такое деление возникло недавно. В 1990 году вышла посвященная этой теме публикация. Деление было сделано на основе последовательности рибосомной РНК. Если раньше для того, чтобы описать какой-либо новый вид, нужно было изучить организм, описать, как он питается, его морфологию, и только после этого его можно было классифицировать, то сейчас классификацию организма можно провести, даже не зная, как он выглядит. Достаточно просеквенировать (определить последовательность нуклеотидов) его рибосомную РНК. И поскольку для многих организмов последовательность рибосомной РНК известна, то по степени сходства этих РНК строится классификация, а не по внешнему сходству или по особенностям метаболизма. Некоторые группы архебактерий так и были описаны: рибосомные РНК есть, а сами организмы пока никто не видел. В чем же смысл перехода к классификации по степени сходства рибосомной РНК? Рибосомная РНК отражает родство по происхождению, в то время, как одинаковая форма может быть у совершенно неродственных животных. Если вспомнить лягушку, крокодила и бегемота, то окажется, что глаза у них из воды торчат похожим образом. Но эти животные относятся к разным классам. То есть, построение классификации на основе рибосомной РНК отображает родство организмов, но зато зачастую не отражает сходство в образе их жизни. Почему выбрана именно рибосомная РНК? Потому что это самая консервативная, т.е. наиболее медленно меняющаяся, часть генома. Ниже на рисунке представлено дерево родства разных организмов. На нем выделяются группы бактерий, архей и эукариот.Эти группы, более высокого ранга, чем царства. Их называют надцарства или домены. Термин домен используется в разных науках. В данном случае, в систематике, «домен» обозначает группу (рангом выше царства), объединяющую разные организмы, обладающие определенным набором общих черт.

Что же общего у бактерий и архей, что отличает их от эукариот?

Строение клетки прокариот
У прокариотических клеток есть цитоплазматическая мембрана, также как и эукариотических. У бактерий мембрана двуслойная (липидный бислой), у архей мембрана довольно часто бывает однослойной. Мембрана архей состоит из веществ, отличных от тех, из которых состоит мембрана бактерий. Поверхность клеток может быть покрыта капсулой, чехлов или слизью. У них могут быть жгутики и ворсинки.
Клеточное ядро, такое как у эукариот, у прокариот отсутствует. ДНК находится внутри клетки, упорядоченно свернутая и поддерживаемая белками. Этот ДНК-белковый комплекс называется нуклеоид. У эубактерий белки, которые поддерживают, ДНК отличаются от гистонов, которые образуют нуклеосомы (у эукариот). А у архибактерий гистоны есть, и этим они похожи на эукариот. Энергетические процессы у прокариотов идут в цитоплазме и на специальных структурах - мезосомах (выростах клеточной мембраны, которые закручены в спираль для увеличения площади поверхности, на которой происходит синтез АТФ). Внутри клетки могут находиться газовые пузырьки, запасные вещества в виде гранул полифосфатов, гранул углеводов, жировых капель. Могут присутствовать включения серы (образующейся, например, в результате бескислородного фотосинтеза). У фотосинтетических бактерий имеются складчатые структуры, называемые тилакоидами, на которых идет фотосинтез. Таким образом, у прокариот, в принципе, имеются те же самые элементы, но без перегородок, без внутренних мембран. Те перегородки, которые имеются, являются выростами клеточной мембраны.
Форма прокариотических клеток не так уж и разнообразна. Круглые клетки называются кокки. Такую форму могут иметь как археи, так и эубактерии. Стрептококки – это кокки, вытянутые в цепочку. Стафилококки – это «грозди» кокков, диплококки –кокки, объединенные по две клетки, тетрады - по четыре, и сарцины – по восемь. Полочкообразные бактерии назыаются бациллами. Две палочки – диплобациллы, вытянутые в цепочку – стрептобациллы. Еще выделяют коринеформные бактерии (с расширением на концах, похожим на булаву), спириллы (длинные завитые клетки), вибрионы (коротенькие загнутые клетки) и спирохеты (завиваются не так, как спириллы). Ниже проиллюстрировано все выше сказанное и приведены два представителя архебактерий.

Хотя и археи, и бактерии относятся к прокариотическим (безядерным) организмам, строение их клеток имеет некоторые существенные отличия. Как уже было отмечено выше, бактерии имеют липидный бислой (когда гидрофобные концы погружены в мембрану, а заряженные головки торчат с двух сторон наружу), а археи могут иметь монослойную мембрану (заряженные головки имеются с двух сторон, а внутри единая целая молекула; эта структура может быть более жесткой, чем бислой). Ниже представлено строение клеточной мембраны архебактерии.

Бактерии и археи отличаются строением и размером РНК-полимеры. В состав бактериальных РНК-полимераз входит 4-8 белковых субъединиц, в сотав эукариотических РНК-полимераз входит 10-14 белковых субъединиц, а у архей размер промежуточный: 5-11 субъединиц. Рибосомы бактерий меньше рибосом эукариот и меньше, чем рибосомы архей (которые также имеют промежуточные размеры).
К прокариотам относятся архебактерии, бактерии и синезеленые водоросли. Прокариоты — одноклеточные организмы, у которых отсутствуют структурно оформленное ядро, мембранные органоиды и митоз.
Размеры — от 1 до 15 мкм. Основные формы: 1) кокки (шаровидные), 2) бациллы (палочковидные), 3) вибрионы (изогнутые в виде запятой), 4) спириллы и спирохеты (спирально закрученные).
1 — кокки; 2 — бациллы; 3 — вибрионы; 4—7 — спириллы и спирохеты.

1 — цитоплазматическая мемб-рана; 2 — клеточ-ная стенка; 3 — слизис-тая кап-сула; 4 — цито-плазма; 5 — хромо-сомная ДНК; 6 — рибосомы; 7 — мезо-сома; 8 — фото-синтети-ческие мемб-раны; 9 — вклю-чения; 10 — жгу-тики; 11 — пили.
Бактериальная клетка ограничена оболочкой. Внутренний слой оболочки представлен цитоплазматической мембраной (1), над которой находится клеточная стенка (2); над клеточной стенкой у многих бактерий — слизистая капсула (3). Строение и функции цитоплазматической мембраны эукариотической и прокариотической клеток не отличаются. Мембрана может образовывать складки, называемые мезосомами (7). Они могут иметь разную форму (мешковидные, трубчатые, пластинчатые и др.).
На поверхности мезосом располагаются ферменты. Клеточная стенка толстая, плотная, жесткая, состоит из муреина (главный компонент) и других органических веществ. Муреин представляет собой правильную сеть из параллельных полисахаридных цепей, сшитых друг с другом короткими белковыми цепочками. В зависимости от особенностей строения клеточной стенки бактерии подразделяются на грамположительные (окрашиваются по Граму) и грамотрицательные (не окрашиваются). У грамотрицательных бактерий стенка тоньше, устроена сложнее и над муреиновым слоем снаружи имеется слой липидов. Внутреннее пространство заполнено цитоплазмой (4).
Генетический материал представлен кольцевыми молекулами ДНК. Эти ДНК можно условно разделить на «хромосомные» и плазмидные. «Хромосомная» ДНК (5) — одна, прикреплена к мембране, содержит несколько тысяч генов, в отличие от хромосомных ДНК эукариот она не линейная, не связана с белками. Зона, в которой расположена эта ДНК, называется нуклеоидом . Плазмиды — внехромосомные генетические элементы. Представляют собой небольшие кольцевые ДНК, не связаны с белками, не прикреплены к мембране, содержат небольшое число генов. Количество плазмид может быть различным. Наиболее изучены плазмиды, несущие информацию об устойчивости к лекарственным препаратам (R-фактор), принимающие участие в половом процессе (F-фактор). Плазмида, способная объединяться с хромосомой, называется эписомой .
В бактериальной клетке отсутствуют все мембранные органоиды, характерные для эукариотической клетки (митохондрии, пластиды, ЭПС, аппарат Гольджи, лизосомы).
В цитоплазме бактерий находятся рибосомы 70S-типа (6) и включения (9). Как правило, рибосомы собраны в полисомы. Каждая рибосома состоит из малой (30S) и большой субъединиц (50S). Функция рибосом: сборка полипептидной цепочки. Включения могут быть представлены глыбками крахмала, гликогена, волютина, липидными каплями.
У многих бактерий имеются жгутики (10) и пили (фимбрии) (11). Жгутики не ограничены мембраной, имеют волнистую форму и состоят из сферических субъединиц белка флагеллина. Эти субъединицы расположены по спирали и образуют полый цилиндр диаметром 10-20 нм. Жгутик прокариот по своей структуре напоминает одну из микротрубочек эукариотического жгутика. Количество и расположение жгутиков может быть различным. Пили — прямые нитевидные структуры на поверхности бактерий. Они тоньше и короче жгутиков. Представляют собой короткие полые цилиндры из белка пилина. Пили служат для прикрепления бактерий к субстрату и друг к другу. Во время конъюгации образуются особые F-пили, по которым осуществляется передача генетического материала от одной бактериальной клетки к другой.
Спорообразование у бактерий — способ переживания неблагоприятных условий. Споры формируются обычно по одной внутри «материнской клетки» и называются эндоспорами. Споры обладают высокой устойчивостью к радиации, экстремальным температурам, высушиванию и другим факторам, вызывающим гибель вегетативных клеток.
Размножение. Бактерии размножаются бесполым способом — делением «материнской клетки» надвое. Перед делением происходит репликация ДНК.
Редко у бактерий наблюдается половой процесс, при котором происходит рекомбинация генетического материала. Следует подчеркнуть, что у бактерий никогда не образуются гаметы, не происходит слияние содержимого клеток, а имеет место передача ДНК от клетки-донора к клетке-реципиенту. Различают три способа передачи ДНК: конъюгация, трансформация, трансдукция.

— однонаправленный перенос F-плазмиды от клетки-донора в клетку-реципиента, контактирующих друг с другом. При этом бактерии соединяются друг с другом особыми F-пилями (F-фимбриями), по каналам которых фрагменты ДНК и переносятся. Конъюгацию можно разбить на следующие этапы: 1) раскручивание F-плазмиды, 2) проникновение одной из цепей F-плазмиды в клетку-реципиента через F-пилю, 3) синтез комплементарной цепи на матрице одноцепочечной ДНК (происходит как в клетке-доноре (F +), так и в клетке-реципиенте (F -)).
Трансформация — однонаправленный перенос фрагментов ДНК от клетки-донора к клетке-реципиенту, не контактирующих друг с другом. При этом клетка-донор или «выделяет» из себя небольшой фрагмент ДНК, или ДНК попадает в окружающую среду после гибели этой клетки. В любом случае ДНК активно поглощается клеткой-реципиентом и встраивается в собственную «хромосому».
Трансдукция — перенос фрагмента ДНК от клетки-донора к клетке-реципиенту с помощью бактериофагов.
Вирусы
Вирусы состоят из нуклеиновой кислоты (ДНК или РНК) и белков, образующих оболочку вокруг этой нуклеиновой кислоты, т.е. представляют собой нуклеопротеидный комплекс. В состав некоторых вирусов входят липиды и углеводы. Вирусы содержат всегда один тип нуклеиновой кислоты — либо ДНК, либо РНК. Причем каждая из нуклеиновых кислот может быть как одноцепочечной, так и двухцепочечной, как линейной, так и кольцевой.
Размеры вирусов — 10-300 нм. Форма вирусов: шаровидная, палочковидная, нитевидная, цилиндрическая и др.
Капсид — оболочка вируса, образована белковыми субъединицами, уложенными определенным образом. Капсид защищает нуклеиновую кислоту вируса от различных воздействий, обеспечивает осаждение вируса на поверхности клетки-хозяина. Суперкапсид характерен для сложноорганизованных вирусов (ВИЧ, вирусы гриппа, герпеса). Возникает во время выхода вируса из клетки-хозяина и представляет собой модифицированный участок ядерной или наружной цитоплазматической мембраны клетки-хозяина.
Если вирус находится внутри клетки-хозяина, то он существует в форме нуклеиновой кислоты. Если вирус находится вне клетки-хозяина, то он представляет собой нуклеопротеидный комплекс, и эта свободная форма существования называется вирионом . Вирусы обладают высокой специфичностью, т.е. они могут использовать для своей жизнедеятельности строго определенный круг хозяев.
В цикле репродукции вируса можно выделить следующие стадии.

- Осаждение на поверхности клетки-хозяина.
- Проникновение вируса в клетку-хозяина (могут попасть в клетку-хозяина путем: а) «инъекции», б) растворения оболочки клетки вирусными ферментами, в) эндоцитоза; попав внутрь клетки вирус переводит ее белок-синтезирующий аппарат под собственный контроль).
- Встраивание вирусной ДНК в ДНК клетки-хозяина (у РНК-содержащих вирусов перед этим происходит обратная транскрипция — синтез ДНК на матрице РНК).
- Транскрипция вирусной РНК.
- Синтез вирусных белков.
- Синтез вирусных нуклеиновых кислот.
- Самосборка и выход из клетки дочерних вирусов. Затем клетка либо погибает, либо продолжает существовать и производить новые поколения вирусных частиц.

Вирус иммунодефицита человека поражает главным образом CD 4 -лимфоциты (хелперы), на поверхности которых есть рецепторы, способные связываться с поверхностным белком ВИЧ. Кроме того, ВИЧ проникает в клетки ЦНС, нейроглии, кишечника. Иммунная система организма человека утрачивает свои защитные свойства и оказывается не в состоянии противостоять возбудителям различных инфекций. Средняя продолжительность жизни инфицированного человека составляет 7-10 лет.
Источником заражения служит только человек — носитель вируса иммунодефицита. СПИД передается половым путем, через кровь и ткани, содержащие вирус иммунодефицита, от матери к плоду.
Перейти к лекции №8 « Ядро. Хромосомы»
Перейти к лекции №10 « Понятие об обмене веществ. Биосинтез белков»